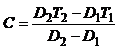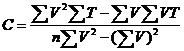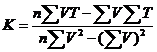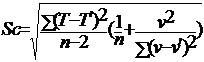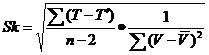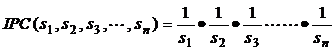|
第五篇 昆虫生态学
第一章昆虫生态学的基本概念
第一节 系统的基本概念
一、什么是“系统”
系统:是许多相互作用又相互联系的物质单元或成分的集合体,它们之间相互依赖又相互制约,成为一个整体。
系统的属性:
1.系统的整体性 系统虽是由许多离散亚系统组成,但它们之间却又集合成为一个具有一定功能的整体。
2.系统的界限和功能 系统都有一定的范围,这种范围的划分,一是根据系统本身所具有的功能,二是根据人们所研究的对象而定。
3.系统的有序性 有序原则体现在系统内部的层次结构。层次原则提示着系统内部各组成分间存在着包含、隶属、支配、服从等传递关系。
4.与环境的融合性 人们常把所研究的对象作为系统的中心,而除此以外的一切均视作环境。
二、系统的反馈机制
反馈现象:系统除受外界环境的输入影响外,还受系统本身的自我控制的影响。也即一个系统在其发展变化过程中可影响其本身的动态或行为。当某一输出状态变量又反过来变为输入变量而影响到状态的动态时,称为反馈现象
正反馈:则为所有输出变量对初始变量的刺激或干扰均有加强的性质。
负反馈:最终输出变量对初始变量的刺激或干扰均有削弱或衰减作用。
第二节 限制因子的原理----生物对生活环境的忍受律和最低定律
一、限制因子的概念
限制因子:当某种或几种基本物质的可利用量最接近于所需要的临界最小量时,这种或这些基本物质便将成为一个限制因子。
二、利比赫的最小因子定律
最小因子定律:当一种或几种限制因子低于需要量的最低阈值时,作物的生长、繁殖或生产将被抑制,而作物的产量直接与这些限制因子的施入量成正比。
最高量定律:当某些因子的存在量高于生物所需要的最高量时,也同样可成为该生物的限制因子。
三、谢尔福德的耐受性定律
耐受性定律:任何一个生态因子在数量或质量上的不足或过多,当接近或达到某种生物的耐受性限度时,就会使该种生物衰退或不能生存。
耐受性定律的特点:1.它不仅考虑到因子量的过少,而且也考虑到因子量的过多。2.不仅考虑到外界因子的限制作用,也考虑到生物本身的耐受能力。3.还考虑到因子之间的相互作用。
第三节 生态平衡及其应用意义
一、生态平衡的概念
生态平衡:是在一定的时间和相对稳定的条件下,生态系统各部分的结构与功能处于相对适应、协调的动态平衡之中。其实质是在特定时间、空间里群体和其所处环境相互关系中种种物质循环和能量转换的相互依赖又相互制约的相对平衡状态。
二、生态平衡的机制
生态平衡的实质是在特定时间、空间里群体和其所处环境相互关系中种种物质循环和能量转换的相互依赖又相互制约的相对平衡状态。
生态系统的物质循环与能量转换,即由不能为动物利用的光能通过植物的吸收转化为动物可以利用的化学能,再通过动植物间营养物质不断地循环,从无机态变为有机态,再由有机态变为无机态,相互不断地转换着,这种能量转换和物质循环,既相互直辖市又相互矛盾,推动着生态系统的变化、发育和演替,同时又保持着相对稳定的动态平衡。动态是指它是在不断变化着,而平衡是指在一定特定时间和空间里又保持着恒定的状态。
所以生态平衡就是在某一特定条件下,生物群体之间以及生物群体与环境系统之间相互依赖、相互制约的关系,而维持着某种恒定的状态。因为生命系统和环境系统都在随时发生变异,但生态系统内具有反馈机制,在遇到外来的干扰或内部的变化时,它有一种反馈或回弹的能力。
生态平衡包括三方面:即结构、功能和输入、输出物质在数量上的平衡。
第二章 有机体与生活环境
第一节 生活环境的类别
在生态学中,环境一般是指除所研究的生物有机体外,周围所有因素的总和。它包括空间以及其中可以直接或间接影响有机体生活和发展的各种因素。
环境因子:
非生物因子(abiotic factors 无机因子)
气候因素:温度、湿度、降水、光、风等
土壤因素:土壤温度、土壤湿度、土壤化学性质、土壤机械组成等。
生物因子(biotic factors同种生物其他有机体或异种生物有机体)
食物因素:食物种类、生长发育状况、质量
天敌因素:天敌昆虫、其他动物、致病微生物等
人为因子:人类在生产实践活动中对昆虫所产生的影响。
史密斯(Smish,1935)人环境因子对于种群数量变动的作用角度,将环境因子分为密度制约因子(density dependent factors)和非密度制约因子(density independent factors)。前者台食物、天敌等生物因子,它们影响大小随种群密度的变化而变化;后者则如气候、土壤等因子,它们的影响大小与种群的密度无关。
第二节有机体与非生物环境
有机体的非生物环境主要是气候因素。气候因素可以直接影响昆虫的生长发育、生存和繁殖,从而造成种群发生期、发生量和为害程度的差异。气候还可以通过对寄主植物或害虫天敌等生物因子的作用,间接地影响害虫的发生。
一种害虫在不同地区或同一地区的不同年份,其发生危害程度,在很大程度上决定于气候条件的适合与否。
气候因素包括温度、湿度、光照、风等,但对昆虫起基本作用的是热和水,即温度和湿度。
温度对昆虫的作用
一、虫热能的获得、散失和调节
1、热能的获得 昆虫属于变温动物,其进行生命活动所需的热能来源,主要是太阳的辐射热,其次是由本身代谢所产生的热能(代谢热),但在很大程度上取决与周围环境的温度。
2、热能的散失 昆虫体积小,表面积大,所以热能散失的主要途径是伴随着水分的蒸发以及向体外传导和辐射热能。
3、热能的调节 改变呼吸强度、水分蒸发强度、改变行为来调节体温。
二、昆虫对温度的适应范围
昆虫只有在一定的温度范围内才能进行正常的生长发育,超过这一范围(过高或过底)其生长发育就会停滞,甚至死亡。因此,根据温度对温带地区昆虫的影响,可将温度划分为五个温区:
(一)、适温范围
1、高适温范围
2、最适温范围
3、低适温范围
(二)、临界致死高温范围
(三)、致死高温范围
(四)、临界致死低温范围
(五)、致死低温范围
三、温度与昆虫发育速度的关系
温度对昆虫发育速度的影响最为显著,平均温度是影响昆虫发生期的重要因素;昆虫的新陈代谢作用是由在各种酶和激素的作用下的一系列生物化学反应组成的,其速度在一定范围内,随着温度的升高而加快。
所以,昆虫的发育速度(V)与温度(T)成正比,而完成发育的时间(D)与温度成反比。
四、 效积温法则
昆虫发育速度与温度之间的关系用有效积温法则来定量表示。
1、有效积温法则的概念
昆虫和其他生物一样,完成其发育阶段(卵、幼虫、蛹、成虫或整个世代)需要从外界获取并积累一定的热能,而完成此一发育阶段所需的总热量是一个常数。积累的总热能以发育历期与此期间的平均温度的乘积表示,称为积温常数(k’)。
即:K ′=DT 式中:K ′代表积温常数,单位为“日温度”,D代表发育历期,T代表发育温度。昆虫是在发育始点(c)以上的温度才开始发育的,所以k=D(T-C),K代表有效积温常数,接下来推导,T =C+K/D
我们取V =1/D(发育速度,单位时间内完成发育的比值)
所以有
T=C+VK K’积温常数
K 有效积温常数D 发育历期T 发育温度V 发育速度C 发育起点
2、测定昆虫发育始点和有效积温的方法
定温法:
在不同温度的温箱内饲养昆虫,观察其在不同定温下的发育历期
⑵、自然变温法:
不需要恒温设备,将供试昆虫分期分批在自然条件下饲养,利用自然界季节性和昼夜的温度变化,而获得在不同日平均的发育历期。试验次数10次以上。
此法,获得的结果比较符合实际情况,但试验所需时间较长。
⑶、人工变温法:
在人工气候室内模拟自然界气温的季节和昼夜变化饲养昆虫,以获得多组不同日平均温度下的发育历期,或在温箱内饲养,作人工级跳式变温处理,即将供试昆虫每天经历6~10小时的较高的温度,而另外14~18小时则给以较低的温度,用加权平均法,求得日平均气温,而获得多组在不同日平均温度下的发育历期。
以上两种方法的温度组合(实验次数n)一般应在5次以上,并保持试虫的适宜湿度和相同的喜食饲料条件。
现举例介绍上述几种测定C和K的方法
2.1、 两种温度组合下求C、K
在第一种温度(T1)与第二种温度(T2)下饲养试虫,分别获得其发育历期D1和D2, 代入K=D(T-C) 得
K=D1(T1-C)
K=D2(T2-C)
解得:
例:在18和25下饲养粘虫卵,其卵期分别为9天和3.5天。则
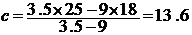 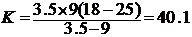
2.2、5种或5种以上温度组合测定K和C
因N>=5,T=C+KV为直线方程,可根据Richardson(1935)的“最小二乘方”求系数的公式求得K和C:
最后用求得的理论值C代入T=C+KV, T=T`±Sc
定温法:求得温度的理论值(T`)和其与温度实测值(T)之差的平方和,计算C和标准差(Sc)
变温法:由于实验取样计算的关系,有一定误差,因此要用以下方法计算C与K的标准差。
故在自然变温下,
3、积温法则的应用
a、预测农业害虫和天敌的地理分布及其在某地一年发生和代数。
b、预测农业害虫与天敌的发生期。
c、利用于初选保存天敌低温和释放天敌适期。
d、利用于选择和引进天敌。
4、有效积温法则在应用上的局限性
a昆虫的发育速度受温度影响外,还受食物和湿度的影响。而此法则只有在温度对该虫的生长发育起主导作用时,才较为准确。
b有效积温法则是以温度与发育速度成直线关系为前提。即在最适温度范围准确而对于高、低适温范围此法反应不出来。
c计算发育起点(C)与有效积温(K)是在定温下得到的数据与自然变温下不同,变温下较恒温下较快。
d 注意昆虫实际环境的小气候
e 昆虫的不同地理种群发育起点也完全不同。
f 无法用某些有滞育阶段昆虫的世代数计算。
五、高、低温对昆虫存活的影响
在季节明显变化的温带地区,高、低温对昆虫存活的影响很大,是昆虫种群数量变动的重要因素之一。
(一)高温与耐热(heat hardiness)
1、高温对昆虫发育的影响
A、高温可抑制昆虫发育,对成虫影响较大。
B、夏季高温出现的强度和延续时间引起死亡率的差异。
2、影响耐热力的因素
A、昆虫种类、生活环境
B、同一种不同发育阶段
3、高温对昆虫致死的原因
A、高温引起体内水分过量蒸发而使昆虫致死
B、高温使体内性蛋白质发生凝结而致死
(二)、低温与耐寒(cold hardiness)
1、过冷却现象和耐寒
2、影响昆虫耐寒力的因素
A、不同发育阶段和龄期
B、昆虫的生理状态不同(含水量、脂肪含量)
C、同一发育阶段不同季节
3、低温致死的原因
A、不很低的温度下致死作用
B、在0℃以下很低温度下致死作用
六、温度对昆虫繁殖的影响
1、影响繁殖的温度主要作用在成虫期,而影响发育的温度主要作用在卵、幼虫、蛹期。
2、昆虫的繁殖力在一定适温范围内随着温度的升高而增大但适合于繁殖的适温范围比适合生长发育的温度范围要小。
总之在较低的温度下,虽然成虫寿命长,但性腺仍不能发育成熟,以致不能交配产卵或产卵量少。在过高的温度下,精子不易形成而失去活力,常常引起不孕,也影响交配产卵活动。在适宜温度下,一般成虫性成熟快,繁殖力大。
第三节 有机体与生物环境间的关系
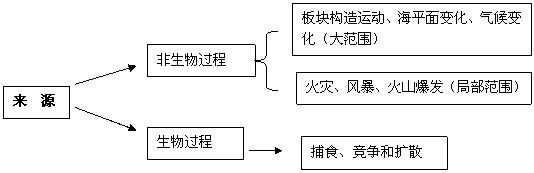
生物因素是环境因素的重要组成部分,它包括环境中的一切生物。生物因素既影响昆虫的生长、发育、繁殖和分布,又影响昆虫种群密度变化。植食性昆虫受寄主植物和天敌的影响尤为明显。同时种间和种内的密度变化在一定条件下也影响种群的盛衰。
一、食物链和食物网
食物链:以植物为起点的彼此依存的食物联系的基本结构。
食物网:错综复杂的食物联系的结构。
二、种间竞争和种内竞争
自然界中生物间存在相互依存和相互制约的复杂关系。一般都是对由食物或居住空间的竞争原因引起的。
种间竞争的关系是生物在长期进化过程中形成的适应性,因而它们的生长发育和繁殖都存在一定的时序性。如同一种作物上,有的害虫出现的较早,有的害虫出现的较晚。种间竞争的适应性沾表现在居住空间方面。种间竞争可表现于食物链的各个环节,竞争的结果往往引起相互的适应。
竞争排斥原理:两个种在竞争过程中各得其所的共同利用环境中的资源。
自然界中种内竞争的影的现象也很普遍。同一空间内任何一种植物或动物的密度超过该空间所能容纳的程度时,经常发生自然淘汰现象而保存物种的延续。在种群密度大而食料不足时,常有自残现象而保存该物种。种内竞争也是物种对环境的适应表现。
三、 生物因素对昆虫的生态效应
生物因素对昆虫的生态效应与非生物因素比较起来,其特点主要表现在以下几方面:
1. 对昆虫种群的影响不均匀性。
2. 与种群密度大小的关系。
3. 昆虫对环境的适应性不同。
4. 相关的物种间互为生物环境,彼此影响对方种群的数量。
第四节 生物对环境的适应
一、昆虫的休眠与滞育
休眠(dormancy):是昆虫种在个体发育过程中对不良外界条件的一种暂时性的适应性。当这种不良的条件一旦消除而能满足其生长发育的要求时,昆虫便可立即停止休眠而继续正常地生长发育。
滞育(diapause):是昆虫种在系统发育过程中本身的生活方式与其外界生存因素间不断矛盾统一的结果,是一种内在的比较稳定的遗传性表现。即使给予适宜的温度或食物条件,也不能阻止滞育的发生。
兼性滞育:不出现在固定世代,随地理、气候、食物等因素而变动。
专性滞育:出现在固定的世代及虫期。
二、昆虫的扩散与迁飞
1.扩散:是昆虫在个体发育中日常的、在小范围内的分散或集中,称为扩散。
扩散类型:
A.完全靠外部因素传播
B.由虫源向外扩散
C.由于趋性所引起的小范围的分散或集中
2.迁飞:指某些昆虫的成虫,在某一时期内从虫源地区成群地、远距离地迁飞到另一地区繁殖为害,从而造成迁入地区严重的虫灾。
3.迁飞昆虫的种群特征与类型
迁飞昆虫的种群特征:
A. 种群数量长期具有季节性“突增”、“突减”现象,上下两代间发生数量十分悬殊。
B. 在一个相当大区域内种群有“同期突发”现象,即大区域内同时突然发生。
C. 种群在上下两代的发育进度不符合。
D. 成虫发生期间雌虫卵巢发育有不连续现象
迁飞昆虫的类型:
A 无固定繁殖基地,连续性迁飞类型
B 有固定繁殖基地的迁飞类型(如蝗虫)
C 越冬越夏迁飞类型(如七星瓢虫)
D 蚜虫迁飞类型(有翅型)
4.迁飞昆虫的种型分化•
迁飞从生物学意义上来是一种行为多态现象,有的个体可作长时间的持续飞行,为“迁飞型”;有的个体只能作短时间的飞行,为“居留型”。种型分化决定于其内在的基因遗传力,也受环境的影响。环境因素包括:光照周期、食料条件的不适宜或缺乏、温度、拥挤度。
第五节 生物对环境的适应---生物钟、行为调节
一、生物钟
1.生物钟的概念
生物钟(biological clocks):生物的生理机能和生活习性受着内在的、具有“时钟”性能的生理机制的控制,这种生理机制称为生物钟。
2.生物钟的类型:
类型1:生物钟的发生与对光的接受器复眼无关,而其对外界光周期的感受直接由脑部的某些细胞组织所控制。
类型11:生物钟主要由光接受器复眼所控制。在完全黑暗或完全光亮条件下失去节律。
二、昆虫基本行为的适应
昆虫有关的行为有四大类型:趋性(taxes)、反射(reflexes)、本能(instincts)、学习(learning)。
趋性:低等动物趋向刺激发源地的行为(趋化、光、色、嫩性等)
反射:通过神经系统对刺激所发生的有规律的反应
本能:即受外界刺激影响又受昆虫体内的特殊环境影响。
学习:通过后天的多次经历或刺激而产和的经验。
第六节 天敌因素对昆虫的影响
天敌因素对昆虫的影响定义:昆虫在生长发育过程中,常由于其它生物的捕食或是寄生作用而死亡,这些生物成为昆虫的天敌。
主要包括:致病微生物、天敌昆虫和食虫动物。
一、致病微生物(简介)
主要从下面四种微生物讲
1.细菌
2.真菌
3.病毒
4.线虫
习惯上呢,也将病原原生动物归于病原微生物(如蝗虫微孢子虫、玉米螟孢子虫等)。
二、天敌昆虫
1.捕食性天敌的特点: 身体一般比猎物大,除了由于适应捕捉和取食猎物的特殊需要,而要在口器、前足或是产卵器上发生变异外,形态上其他方面较少发生变化,成虫、幼虫搜索猎物的目的是为了取食,都是猎物体外行自由生活,除个别例外(如芫箐、一些捕食性的灰蝶幼虫),成虫和幼虫都为捕食性,有的捕食同一种猎物,通常需要捕食许多猎物才能完成其发育,猎物被杀死破坏的速度快。
2.捕食者与猎物相互关系的模型:
主要介绍霍林圆盘方程(holling,1959)和哈塞尔(hassell,1969)干扰作用两种模型。
①功能反应是表示在不同的猎物密度下,每个捕食者与被捕食者(猎物)之间的数量关系。多数无脊椎动物(包括昆虫)的功能反应呈负加速曲线,是逆密度制约的。所以其数学模型可以用此方程表示:即圆盘方程(霍林圆盘方程)。
Na=(a’*N*T)/(1+Th*a’*N)简化
1/Na=(1/ a’*T)*(1/N)*(Th/T)
令1/ a’=B Th= A T=1(1昼夜)1/ Na=A+B/N
用最小二乘方可求得A、B。Th即A,a’即1/B。当N趋向于无穷时,Na即为1/A,这是捕食者最大捕食量(饱和捕食量)N 为猎物的密度
Na为被捕食的猎物数量
Th 为捕食者可以用以发现猎物的时间
a’ 为瞬间攻击率(功能系数),
T 为处理时间
②干扰作用(在一定的空间内,捕食者自身的数量对其捕食的猎物数量有着干扰作用。哈塞尔(1969))
E=Q*P-m LgE=LgQ – M*LgP
E为捕食量E=Na/(N*P) Q为搜索常数
m为干扰常数 p为一定时间内捕食者的数量
Na/p为1头捕食者平均捕食猎物的数量
N为猎物密度(预设)
3.寄生性天敌昆虫
特点:身体一般比寄主小,幼虫期因不需要寻找寄主,足、眼都退化,成虫搜索寄主,主要是为了产卵,一般不杀死寄主,除成虫自由生活外,其他发育阶段或至少幼虫在寄主体内或体表生活,即不能离开寄主独立生活,与寄主关系密切,成虫与幼虫食性不同,幼虫为肉食性,在一个寄主上可发育成一个或多个个体,寄主被寄生后致死作用缓慢。
寄生性天敌昆虫的寄生类型
1.根据寄生性昆虫的幼虫在寄主体内或体表取食、完成生长发育的情况,可以分为内寄生和外寄生
2.根据寄主内寄生性昆虫的种类的多少可分为:独寄生和共寄生
3.根据一个寄主内寄生的同一种寄生性昆虫的数量,可分为单寄生、多寄生
4.根据寄生性昆虫在寄主体内完成发育情况,可分为完寄生和过寄生
5.根据寄生性昆虫与寄主的寄生关系是直接还是间接可分为原寄生或是重寄生
6.根据寄生性天敌对寄主种类选择范围的大小分为单择性、寡择性和多择性
三、天敌昆虫与农业害虫间的信息联系
生物与生物之间的基本联系是营养联系,同时也是化学信息联系,以保证营养联系的完成。天敌昆虫与猎物(或寄主)之间也有着化学信息联系,以保证搜寻猎物(或寄主)和猎物受到天敌侵扰时向同伴发出警告。在这里呢,性信息化合物分为两类。
1.信息素(pheromone):一种由昆虫分泌并释放到体外,以引起同种昆虫其他个体产生行为反应的化学物质。主要是性信息素(sexpheromone)、集结信息素(aggregation)、踪迹信息素(trail-marking pheromone)、告警信息素(alarm pheromone)。
2.交互化合物:一种昆虫分泌,并释放到体外,引起另一种昆虫产生行为反应的化学物质。利己素(allomone)、利他素(kairomone)、协同素(synomone)。
四、其他的食虫动物
包括鸟,蜘蛛、树蛙等等。
我们了解一下蜘蛛:通过各地调查,农林中常见种类约百余种。在一块田中的优势种类虽然不多,但个体数量数万或一、二十万只,约占捕食性天敌总数的50—80%。所以,近年来引起了国内外从事植保和有关科研工作人员的注意。对它们的分类、生态、生理、毒理以及对蜘蛛控制和保护利用更做了大量的调查和研究工作。几年来,我国先后发表了许多研究报告,并把蜘蛛编入了天敌图册,得到了应有的重视。经过大面积的综合防治试验,证明通过充分发挥这类天敌的作用,可以大大降低化学农药的用量,由此,减少了生产成本、农药在作物中的残留量及对偶环境的污染,同时也保护了害虫的其他天敌。
第二章 种群生态学
第一节 种群的基本特性与种群结构
一、种群的概念、特点及其分类
1、 种群(POPULATION):是指在一定的生活环境内,占有一定空间的同种个体的总和,是种在自然界存在的基本单位。
2、 种群的特点:
a.种群是通过种内关系组成的一个有机的统一群体,它除具有种的一般生物学属性(形态结构、生活方式、遗传性相同,以及与其它种存在严格的生殖上隔离)外,
b.还具有群体自身的生物学特性(出生率、死亡率、性比、平均寿命、年龄组成、基因频率、繁殖速率、密度及数量变动、空间分布、迁移率、滞育率)。
C.种群是一个自动调节系统,具有群体的信息传递、行为适应、数量反馈控制的功能。
3、 种群的分类:
a.同种昆虫由于长期的地理隔离而形成的种群,称为地理种群(地理亚种,地理宗)。
b.因寄主食物的不同而形成的不同种群,称为食物种群(食物宗、寄主宗)。
c.有时由于需要研究两种或两种以上的种群,称为混合种群。
二 、 种群的结构
1、昆虫种群的结构:即昆虫种群的组成,是指种群内某些生物学特性、对环境适应能力或形态上可以是完全相同的个体群在总体中所占的比例。其中主要是性比和年龄组配
性比:是指成虫或蛹雌性与雄性之比。或以雌虫率表示。
年龄组配:是指一个自然种群中各发育阶段(卵、幼虫、蛹、成虫)和同一发育阶段不同发育进度(如各龄幼虫、各级卵、蛹或成虫)的数量比例或所占的百分率
第二节 种群的空间分布类型
一、种群分布型的概念
种群是指在一定空间内同种动物(或植物)有机组合的群体。任何一个动物种群总要占据一定的生殖场所,拥有一定的食物供应范围和活动领域,即生态学中所谓的“空间”。
种群动态是指任何一个种群在一定空间内适应环境的历史。种群动态一方面表现为数量上的增减;另一方面表现为数量上的波动,必然关系到空间的扩张和收缩。因此衡量一个种群昌盛与否,通常采用两个指标,即数量与空间。
由于种群栖息地内生物的(种性、种内和种间关系)和非生物的(气候、地形、土质等)相互作用的关系,就造成了种群在一定空间内个体扩散分布的一定形式,这种形式就称为种群的空间格局或空间分布型。
二、种群空间分布型的类型
(一)分类依据
种群空间格局的变化不仅随着不同的物种而有差异,而且亦随着同一物种的不同世代,同一世代的不同发育阶段,以及同一发育阶段的不同龄期、不同密度而不同。若物种个体间呈现相互吸引,为聚集分布(aggregated distribution);若个体间相互独立,则为随机分布(random distribution);若个体间相互排斥,则为均匀分布(uniform distribution)。
(二)分类类型
根据种群内个体的聚集程度和方式不同,可把昆虫种群的空间格局分为随机型、均匀型和聚集型三大类。在随机型中包括泊松分布,均匀分布中包括正二项分布,在聚集分布型中包括核心分布(奈曼分布)和负二项分布(嵌纹分布)。
1、 随机型 泊松分布
2、均匀型 正二项分布
3、聚集型
嵌纹分布(负二项分布) 核心分布(奈曼分布)
三、种群个体的离散频次分布方法
1、 随机型
种群内个体独立地、随机地分配到可利用的单位(生物资源)中去,每个个体占据空间任意一点的概率是相等的,即种群内的个体,相互之间是独立的,一个个体的存在位置不影响其它个体存在的位置。
正二项分布(binomial distribution)又叫二项分布、均匀分布或一致格局。所谓正二项分布就是指数为正的二项式展开后所得到的各项分布。
正二项分布的特点是:
1、种群内的个体在空间的散布是均匀的;
2、种群内的个体在空间的分布比较稀疏,不聚集;
3、个体间相互独立,无影响;
4、当调查单位内实查的数值比较大时(即密度大时)可成一个对称的或近似对称的次数分布曲线。
2、均匀型
泊松分布(poisson distribution)
该分布的特点是:
1、种群内个体在空间的分布是比较稀疏的;
 2、种群内个体间是相互独立的,即任一个体在某一抽样单位中出现与否与其它个体是否在该抽样单位中存在无关; 2、种群内个体间是相互独立的,即任一个体在某一抽样单位中出现与否与其它个体是否在该抽样单位中存在无关;
3、其S2/ =1~1.5;
4、当调查单位内实查数值较小时(虫口密度较小时),可得到一条向右倾斜的偏态次数分布曲线,但当调查单位内实查数值增大时,一般指x≥16时,可趋于正二项分布。
3、聚集型
聚集分布是指昆虫个体,因种种原因造成的分布不随机性,这种不随机性的显著特点是疏密不匀。也分两种类型:嵌纹分布、核心分布。
①嵌纹分布(Mosaic distribution)或负二项分布(Negative binomial distribution) 这是昆虫最广泛的一种分布。
该种分布的特点是:
1、由于种群内个体间具有明显的聚集现象或由于环境条件的不均匀性,使种群个体呈现为疏密相嵌,很不均匀的分布。如麦蚜、棉蚜等;
2、种群内各个体在抽样单位中出现的机会不相等;
3、S2/ 通常介于1.5~3.0之间;
4、种群内个体群之间为Poisson分布,而个体群内个体的分布为对数分布。
②、核心分布(contagious distribution)或奈曼分布(Neyman distribution)
该分布的特点是:
1、昆虫种群内的个体在栖息地里呈多数小集团,形成很多核心,这些核心大小大致相等;
2、在每一个抽样单位中,各个体群之间呈泊松分布,而个体群内的个体亦呈泊松分布;
3、种群内个体彼此间不是相互独立的;
4、S2/ =1.5~3.0, 多数昆虫所产卵块孵化为幼虫后,自核心作放射状蔓延,属此种分布。
三、种群空间分布型的测定方法
(一)频次分布测定法
频次分布测定法(determination method of frequency distribution)是一种经典的方法。基本原理是将实查的种群空间分布信息(如均数、方差和取样数),首先整理调查的原始数据,编制实查频次表,然后计算相应的参数,进而依各理论分布公式算出理论频次分布,最后用实际调查整理的频次与对应的所计算理论频次值进行卡方检验,根据其吻合程度来判断该资料属那种分布型,吻合程度是通过查表与卡方值相比而得出的。
频次分布测定的具体步骤如下:
1、 确定调查对象。
2、 选好调查标准地。根据害虫发生的情况和危害程度,选择具有代表性的试验地。
3、 确定调查方法。包括抽样方案的制定、抽样单位的选择和理论抽样数的确定。
4、 整理调查结果。列出每个样方的虫口数(X),和实测频次(f)所组成的频次分布统计表,以求出样本方差(S2)和平均数(x)
5、 按照各分布型的概率通式,计算各项理论概率及其相应的理论次数。
进行卡方检验,测定其实测频次与理论频次之间的差异是否显著。
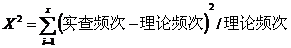 计算卡方值的公式为: 计算卡方值的公式为:
然后根据自由度(f)和概率水平(P)查x2值。为了查表方便,这里提早说明各种空间格局类型条件下的自由度:正二项分布和泊松分布为(n-2),负二项分布和奈曼分布均为(n-3)。在相应和自由度下凡算得卡方累计值大于该自由度下P0.05时的卡方值,则其P〈0.05。表示理论分布与实际分布相符合,也就是不属于该分布。反之,当算的卡方值的P〉0.05时,即表示二者相符合,可以判断为属于该种分布型。
(二)、聚集指标法
聚集指标法(Indies of aggregation)的特点:计算简便,可根据其指标的大小来判断种群的分布类型,并对种群个体的行为,种群扩散的序列变化也能提供一定的信息。但其缺陷是判断分布型比较粗放,只分大类,不及经典频次法具体。具体包括: 扩散系数(C)、 负二项分布K值法、扩散型指数(Iδ)、平均拥挤度、平均拥挤度(X*)与平均数的回归关系法、Taylor幂法
1、 扩散系数(C)
所谓扩散系数就是利用均数和方差是否相等的这一特性,进一步求其S2/ X的比值,用来估计所检验的昆虫种群是否偏离随机型的一个系数,常记为C,其公式为:
C=∑(xi-X )2/(n-1)= S2/X
式中:xi为每个样本内的虫口数;
为虫口平均密度;S2为方差;n为抽样数。
简单判别:C=1时,个体为泊松分布;
C>1时,个体为聚集分布;
C<1时,个体为均匀分布。
2、 负二项分x布K值法
负二项分布中的K值的生物学意义是,K值可以作为种群聚集程度的一个度量:K值的大小,反映聚集度的大小,K值愈大,聚集度愈小,K值愈小,聚集度愈大。如K值趋于无穷大时(一般为8以上时),则逼近poisson分布。
注意:用K值法时,由于K值受抽样单位大小的影响,一般最好在同样大小的抽样样方下进行。
用Ca,即1/K来表示,比较方便。
当Ca=0时,为随机分布;
当Ca>0时,为聚集分布;
当Ca<0时。为正二项分布。K=x2/(s2-x)
3、扩散型指数(Iδ)
扩散型指数(Iδ)最大优点是分布型、抽样数、抽样单位大小独立。
扩散型指数(Iδ)公式:
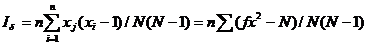
式中:n为抽样数;N为总虫数;xi为第i个样本中的虫口数。
Iδ=1 为poisson分布;
Iδ>1为聚集分布;
Iδ <1均匀分布。
4、平均拥挤度
平均拥挤度是指平均在一个样方内每个个体的拥挤程度,强调的是个体的平均,而不是象x平方那样是样方的平均。
特点:在种群密度较低时,“0”样方很多,虽然平均数X很小,但实际上在集团中的个体间仍极拥挤,种内竞争激烈,因此平均数X很难反应生物因素的相互影响效应,而x﹡可比较真实的反应出种内竞争等生物因素的作用,在生态学中具有很重要的意义。
计算公式为:
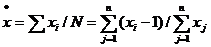
式中K为负二项分布的K值。(大量抽样)
Lwan(1971)提出,用平均拥挤度(X*)与平均数(X)之比值,作为判断种群空间格局的指标。
当X*/ X =1时,为POISSON分布 当X*/ X <1时,为均匀分布
当X*/ X >1时,为聚集分布
5、平均拥挤度()与平均数的回归关系法
Lwao提出用X*与X之间的直线回归关系式中的两个回归系数α,β作为判断种群空间分布型的指标。
X*=α+β α β分别为截距和斜率。
α说明分布的基本成份,β说明基本成分的空间分布型。
α>0时,个体间相互吸引,分布的基本成分的个体群。
α=0时,分布的基本成分是单个个体。
α<0时,个体间相互排斥。
Β=1随机分布,Β>1聚集分布,Β<1均匀分布
而α与β的不同组合,亦提供种群的不同分布类型的信息。
当α=0,β=1时,种群分布型为随机。
(1)α.>0,β=1时, 核心分布、泊松分布-正二项分布
(2)α=0,β>1时, 均为聚集分布 负二项分布
(3)α>0,β>1时 负二项分布
6、Taylor幂法
Taylor指出:在自然种群中,多属不随机分布,样本均值(x)与方差(S2)之间是不独立的,方差常随均值的增加而增加。他从大量的生物学资料中,得出方差的对数值与均数的对数值存在着下列回归关系:
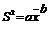 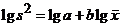
截距a是一个取样统计因素
斜率b是表示当平均密度增加时,方差的增长率,因而它是聚集度对密度依赖性的一个尺度。
最后利用参数a、b的取值,可对种群个体数散布的随机或非随机状况作出判断。方法如下:
6、Taylor幂法为:
对数转换为:
利用参数的取值,可对种群个体散布的随机或非随机状况作出判断。
判断法则如下:
(1)若loga=0,(即a=1),且b=1,则为随机分布;
(2)若loga>0,(即a>1),且b=1,则种群在一切密度下 都是聚集的,但其聚集强度不因种群的改变而变化;
(3)若loga>0,(即a>1),且b>1,则种群在一切密度下都有是聚集的,聚集强度随种群密度的升高而增加;
(4)若loga<0,(即0<a<1),且b<1,则种群密度越高,分布越均匀。
第三节 昆虫种群的数量变动
一、数量变动的参数
昆虫种群数量的变动主要取决于种群基数、繁殖速率、死亡率、和迁移率等。
1、种群基数(N)是指前一代或前一时期某一发育阶段(卵、幼虫、蛹、成虫)在一定空间的平均数量。是估测其下一代或后一时期种群数量变动的基础数据。在调查N的过程中,应注意取样调查的准确性和代表性。
对一些扩散能力强或具有迁飞性的昆虫的成虫,常以1支黑光灯诱集的上代总数量作为下代的种群基数。
也可在一定空间内,标记(如用喷涂颜料、示踪原子等方法)释放和捕回成虫。按释放或捕回的数量比来估计种群基数。
种群基数=(捕回成虫总量/捕回标记成虫量)*释放标记成虫量。
2、繁殖速率(R)
是指一种昆虫种群在单位时间内增长的个体数量的最高理论倍数。它反映了种群个体数量增加的能力。
繁殖率的大小取决于种群的生殖力、性比和一年发生代数。可以用下式表示:
R=(e*f/(m+f))N
其中e为单雌平均生殖力(产卵量),m为雄虫数,f为雌虫数,N为一年发生代数。
3、死亡率和存活率
种群的死亡率(d)是指在一定环境条件下和一定时间内的生态死亡率,它是因时间、环境条件而变化的。一般用在一定时间内种群死亡个体数占总数的百分率表示。存活率s=1-d
4、迁移率 迁移率(M)是指种群在一定时间内迁出个体和迁入个体数量差占总体的百分率。一般情况下,种群无明显的扩散和迁移,其迁移率可视为零。
综上所述,昆虫种群的数量变动的基本模式可以概括为:
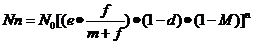
或 Nn=N0〔R×(1-d)×(1-M)〕n
5、昆虫的种群数量变动也可用种群消长指数
(In)表示。消长指数是指种群至第n代(或第n个发育阶段)时的数量与种群起始数量之比。
即: In=Nn/N0=〔e×f/(f+m)×(1-d)×(1-M)〕n
如I=1,表示该种群经N代后,数量未变;
若I>1或I<1,则表示该种群经几代后,其数量增加或下降。
由于昆虫各代的生殖力、性比、死亡率等不同,同一世代的各个发育阶段(卵、各龄幼虫、蛹、成虫)的死亡率等也不相同,这样有时需要计算综合消长指数(I),即:
I=I1×I2×I3……In
I=N1/N0*N2/N1*N3/N2……Nn/Nn-1
二、 种群的生长型和季节消长类型
(一)、 种群的生长型
种群的生长型(Population growth form)又称为“种群在时间上的分布”它是在一定条件下,单种种群在时间序列上数量增减的变化形式。
在实验条件下,它主要有两种类型:
J型生长型,即种群数量开始时增加迅速,后因环境因素抑制,数量突然下降。
S型生长型,即种群数量开始时增加缓慢,随即迅速增加,几呈直线上升,然后又增加缓慢,最终达饱和状态,种群数量不再增加。
但这两种生长型也常因物种和环境而发生改变或联合。
(二)、 种群数量的季节消长类型
昆虫的种群数量是随季节的变化而消长波动的,这种波动在一定空间内常有相对的稳定性,因而形成了昆虫种群的季节消长类型。现将一些重要的农业害虫种群数量的季节消长归纳为以3个类型
1、 单峰型 单峰型是指一年内昆虫种群数量只出现一次高峰。又可分为前峰型和中峰型两类:前者是在生长季节前出现种群数量高峰。后者是在生长季节中期出现高峰。
2、 双峰型 双峰型是在生长季节前、后期(春、秋)各出现一次高峰,故又称为马鞍型。
3、 多峰型 多峰型是指昆虫种群逐季递增。出现多次峰期,故又称为阶梯上升型或波浪型。
第四节 昆虫种群生命表
一、 命表的概念、特点、应用
1、生命表的概念
生命表(life table)是指将一定种群的死亡数量、死亡原因、死亡年龄(时间)等资料列成表,以便分析该种昆虫种群生活史过程中引起数量变动的原因。是研究昆虫数量动态的重要方法之一。
2、生命表的特点
①、系统性:表现在它系统地记录了自然条件下和实验条件下昆虫种群在整个世代从开始至结尾的生存或生殖的情况。
②、阶段性:表现在它分阶段地记录了各虫态,各年龄或各年龄组的生存和生殖情况。
③、综合性:表现在它记录了影响种群数量消长的各种生物和非生物因素的作用形式。
④、关键性:表现在通过对关键因子的分析,找出在错综复杂的综合因素中的关键因子和关键虫期。
3、生命表的应用
①、应用于害虫的数量测报(由描述性的定性的到解释性的定量化的转变)。
②、 应用于评价各种防治措施对控制害虫数量的作用。除了能精确分析出单项措施在某个特定时刻的防治效果外,还能从整个种群数量变动的估值来评价防治措施的最终效果。
③、 应用于害虫的科学管理上。
④、 应用于种群数量变动和模式化表达。即模型的建立。
二、生命表的类型及基本形式
生命表有特定时间生命表(time-specific life table)和特定的年龄生命表(age- specific life table)两种类型,分别适用于世代重叠和世代较离散的昆虫。
特定时间生命表特定时间生命表是指在昆虫种群是静止(后一时间的种群与前一时间的种群的比基本上为1),而世代重叠、年龄组配稳定的前提下,在特定的时间单位内(如月、旬、周、日等)的一种生命表。
它可获得在特定时间内种群的存活率和出生率,适用于世代和年龄组配重叠的昆虫,可估算出种群在各时间内的死亡率、平均生命期望值和世代平均时间,但不能分析死亡原因和关键因素。
1、存活生命表
在特定时间内对种群随机抽样,检查各期的个体数,其个体数差即死亡数,推算各期的死亡率和平均生命期望值。如下表即为假设的存活生命表
下面是所建生命表中各项含义:
x为按年龄或一定时间划分的单位时间期限(如日、周);
Lx为第x天存活的数量;
dx为第x天死亡数量,即lx-Lx+1;
1000qx为第x天时间内死亡率×1000,即为(dx/Lx)×1000;
Lx为相当于时间第x到第x+1时的生存个体平均数,即为(Lx +Lx+1)/2;
Tx为Lx、栏从底层向上累加的共计数,即Tx=Lx+Lx+1,即剩余总寿命;
ex为平均生命期望值(x时间后的个体平均寿命),ex=Tx/Lx,如ex为该虫x1时间的理论平均寿命。
表2.1 假设某虫的存活生命表
下面是横列中各项的计算推导方法:
生命表中只有lx和dx是实际观测值,其他各栏都是统计数值。
Lx+1=lx-dx 如14=13-d3=500-200=300
Lx=(lx+lx+1)/2 or(Lx=lx-1/2dx) L2=700-200/2=600
Lx=Lx+Lx+1+Lx+2 T3=L3+L4+L5+L6+L7+L8=400+200+75+35+15+5=730
Ex=Tx/lx e1=2180/1000=2.18 e4=330/300=1.1
1000qx=dx/lx*1000 1000q1=300/1000*1000=300
2、特定年龄生命表
特定年龄生命表是以种群年龄作为划分时间的标准,系统观察并记载不同发育阶段或年龄区组中的死亡数量,死亡原因以及成阶段的繁殖数量。
表2.3一个理想种群生命表(仿WOODS)
|
年龄(虫期) |
存活数lx |
死亡原因dxF |
死亡数dx |
死亡率100qx (%) |
存活率Sx |
|
卵(N1) |
6000 |
寄生 |
3000 |
50 |
0.5 |
|
一龄幼虫 |
3000 |
天气 |
2000 |
67 |
0.33 |
|
二龄幼虫 |
1000 |
寄生
捕食
小计 |
200
300
500 |
20
30
50 |
0.5
|
|
蛹 |
500 |
寄生或捕食 |
100 |
20 |
0.8 |
|
成虫 |
400 |
性比
(雌虫占40%) |
80 |
20 |
0.8 |
|
雌蛾×2 |
320 |
生育力下降 |
160 |
50 |
0.5 |
|
正常雌蛾×2 |
160 |
成虫扩散与死亡 |
140 |
88 |
0.12 |
期望卵量=正常雌蛾数×每雌蛾的最大产卵量=160/2×200=16000
实际卵量(N2)=20/2×200=2000
趋势指数=N2/N1=2000/6000=0.33
qx为100dx/lx
趋势指数:种群数量的增长倍数
下表为吴坤君等1977年在江西省观察和组建的第四代棉铃虫在棉田的自然种群生命表
表2.4 第四代棉铃虫自然种群生命表
|
发育期(X) |
每一虫期开始时存活数(Lx) |
死亡原因
(dxF) |
每一虫期内死亡数(dx) |
死亡率(qx)(%) |
存活率(sx)
(%) |
|
卵期 |
1000.0 |
捕食性天敌
寄生性天敌
自然损失(未授精等)
合计 |
106.5
76.5
74.5
257.5 |
10.65
7.65
7.45
25.75 |
74.25 |
|
孵代期 |
742.5 |
捕食性天敌
胚胎死亡
自然损失
合计 |
60.1
27.8
352.7
440.6 |
8.10
3.75
47.50
59.35 |
40.65 |
|
1~3龄幼虫期 |
301.8 |
捕食性天敌
寄生性天敌
自然死亡
合计 |
137.2
6.7
40.3
184.2 |
45.43
2.23
13.34
61.00 |
39.00 |
|
4~6龄幼虫期 |
117.7 |
捕食性天敌
寄生性天敌
合计 |
39.8
0.2
40.0 |
33.80
0.20
34.00 |
66.00 |
|
蛹期 |
77.7 |
灌溉等 |
32.6 |
42.00 |
58.00 |
|
成虫
♀×2 |
45.1
44.2 |
性比(♀∶♂=49∶51) |
0.9 |
2.00 |
98.00 |
|
全世代 |
|
|
955.8 |
95.58 |
4.20 |
以预测第五代棉铃虫卵量及其种群增长趋势。
三、种群生命表的分析
1、关键虫期和关键因素的判断
关键虫期和关键因素是指某一虫期和某一因素能极大的影响昆虫整个种群未来数量变化的虫期和因素。
进行关键虫期的分析,至少要有5年或5年以上的同代次的生命表资料,才能进行合理的变量分析。常用的方法有K值图解法和相关回归分析法。
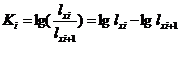 (1)、图解法 K值是前、后相邻的两个发育阶段(或因素)的存活虫数(lx)的比值的对数值。即: (1)、图解法 K值是前、后相邻的两个发育阶段(或因素)的存活虫数(lx)的比值的对数值。即:
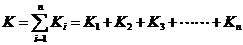 全世代各发育阶段的K值之和,称为K(总K值),即: 全世代各发育阶段的K值之和,称为K(总K值),即:
以年份为横坐标,以K值为纵坐标,绘制总K值和各发育阶段(或各因素)(KI)的坐标图,看那一发育阶段(或因素)的图象与总K值的图象最为相似,则这一发育阶段(或因素)即为关键虫期(或关键因素)。
(2)、相关回归分析法
其中又有斜率(b)法和决定系数(r2)法两种。
①、斜率法以各发育阶段(或各因素)的KI值为自变量(X),全世代总K值为因变量(Y);或以各发育阶段(或各因素)的存活率(或死亡率)的对数值为自变量(X),以下代虫数(或种群增长指数I)的对数值为因变量(Y),代入下式,分别求得各自的b值,
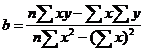
当b>1时,b值最大的发育阶段(或因素)为关系虫期(或关系因素);当b<1时则反之。
②、决定系数法
同上,将各发育阶段(或因素)的自变量(x)和因变量(y),代入下式,分别求得各发育阶段(或各因素)的决定系数(r2),即相关系数(r)的平方。其中发育阶段(或因素)的值最大者,即为关键虫期(或关键因素)。
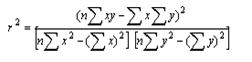
2、种群控制指数庞雄飞(1990)提出利用种群的控制指数(index of population control,IPC)来分析种群数量动态。控制指数是指以被因素作用的种群趋势指数(I′)与原有种群趋势指数(I)的比值,即IPC= I′/I,故IPC值是引起种群趋势指数改变的倍数。趋势指数即增长指数,I=N1/N0。
控制指数是分析控制因素对种群系统控制作用的一个指标。如作用因素I的控制指数为:IPC=I=1/Si;如作用因素为n个,则控制指数为:
种群控制指数的测定,也可用于综合分析天敌、药剂防治等对种群数量发展的影响程度,以评价不同防治措施的效果。
3、种群的存活曲线
⑴ 种群的存活曲线定义:
在某一特定时刻,种群中的同龄个体随时间移动而减少的现象,可以用一条曲线来表示,这条曲线称为存活曲线。如以发育阶段(年龄)为横坐标,存活数(或存活率)为纵坐标,绘成的不同发育阶段的存活数(Lx)曲线。存活曲线是建立预测预报的基础。
⑵ 存活曲线的类型
DEEVEY(1950)比较了各种动物的生命表,指出存活曲线大致分为三种类型
Ⅰ型:这类动物中,绝大多数个体均能实现其平均寿命,待达到其固有的寿命时,几乎同时死亡,就是说,死亡率主要发生在年老的个体。这样Lx曲线呈现明显的上拱形。当存活曲线为该类型时,生命期望ex将随年龄的增加而增加。饥饿的果蝇。如图第一种类型。
Ⅱ型:这类动物在各年龄组均维持同样的死亡率,亦即每个单位时间内或年龄组内死亡数相等,因此Lx曲线呈一直线,也即ex呈常数。例如水螅
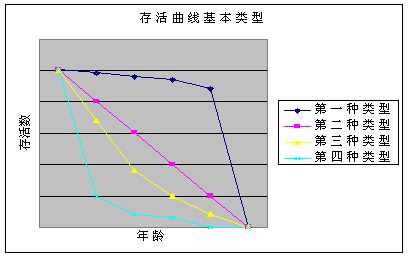 算术直线型,如图第二种类型,即每单位时间内的死亡数为常数。 算术直线型,如图第二种类型,即每单位时间内的死亡数为常数。
对数直线型,如图第三种类型,即每单位时间内的死亡率为常数。
Ⅲ型:这类动物在幼年有极高的死亡率,因此Lx形曲线呈下拱形,这时ex将随年龄增加而减小。如图第四种类型。
存活曲线的意义:了解农业害虫的存活曲线,可明了其易遭伤亡的发育阶段,以分析确定适宜的防治时期。
第五节 昆虫种群的生态对策
一、生态对策
1、生态对策定义------生态对策(bionomic strategy)又称生活史对策(life history strategy):即昆虫在进化过程中,经自然选择获得的对不同生境的适应方式。昆虫的生态对策是其对生态环境适应能力的体现。
2、生态对策的类型
昆虫种群的大小和变化速度主要取决于昆虫种群的内禀增长率(r)和环境容量(k)。种群的内禀增长力是指在特定的环境条件下,具有稳定年龄组配的种群的最大瞬间增长速率。环境容量是指在食物、天敌等各种环境因素的制约,种群可能达到最大的稳定的数量。r反映了昆虫种群的增长速率,k反映了昆虫种群发展的最大范围。所以,当k值保持一定时,r值愈大,种群增长速率愈快,种群愈不稳定;当r值保持一定时,k值愈大,种群发展的范围愈大,种群愈趋向稳定。
根据r值和k值的大小,可将昆虫种群基本上分为两个生态对策类型。
(一)、K—对策者类型r值较小,k值较大;种群密度比较稳定,经常处于环境容量水平,称为K—选择,属于K选择的生物,称为K类有机体。这类有机体体型常较大,寿命与世代亦较长,繁殖能力小,但常具有完善的保护后代的机制,因此,其后代的死亡率较低,通常不具有较强的扩散能力。
(二)R—对策者类型r值较大,k值相应较小;种群密度比较稳定,很少达到环境容量水平,称为R—对策。属于R选择的生物,称为R类有机体,这类生物有机体的体型往往比较小,
寿命以及每个世代的周期较短,繁殖能力很强,但常没有完善的保护后代的机制,因此,其后代的死亡率较高,通常具有较强的扩散迁移能力。
根据K对策和R对策型的特点,可为害虫的防治提供参考。对R类对策的害虫其繁殖力较大,大发生频率高,种群恢复能力强,许多种类扩散迁移能力强,常为暴发性害虫,虽有天敌侵袭,但在其大发生之前控制作用比较小。故对此类害虫防治策略应采取以农业防治为基础,化学防治与生物防治并重的综合防治。单纯的化学防治则由于此类昆虫的繁殖能力强、种群易于在短期内迅速恢复,特别是容易产生抗药性,但在大发生的情况下,化学防治可迅速压低种群数量。同时应研究保护利用和释放相应的天敌昆虫,充分发挥生物防治的控制效应。
中间型:实际上生物的生态对策从K-对策到R-对策是一个连续的系统,称为R-K连续系统。在这个系统中,按照K类选择和R类选择的不同程度排列各种各样的生物,除极端的K对策型和极端的R对策型外,存有许多过渡的中间型。所以这两种对策型的划分也是相对的。
表4.1 R—对策及K—对策的比较表
|
特征 |
R—对策者 |
K—对策者 |
|
气候 |
多变,不确定 |
稳定 |
|
死亡率 |
非密度制约型,常为灾难型 |
密度制约型 |
|
存活率 |
Deevey存活曲线常为Ⅲ型 |
常为Ⅰ、Ⅱ型 |
|
种群密度 |
多变的不稳定的,不饱和的部分常为生态真空,每年去占领 |
稳定、饱和、平衡 |
|
种内竞争 |
变动较大,通常不紧张 |
经常种于紧张状态 |
|
选择利于 |
体形小,发育快,提早繁殖,单次生殖 |
体形较大,发育缓慢,延迟生殖,多次生殖 |
|
寿命 |
短,常少于一年 |
长,常大于一年 |
|
导致 |
提高生产率 |
提高效率 |
二、栖境特性和生态对策的关系
栖境:对任何一种动物来说可以定义为整个生活期间活动所到达的地区。
与生态对策有关的栖境特性可以包括以下三个方面。
1.栖境的稳定性 即在一特定地理位置上,特定生境类型所保持的时间长度。其稳定的意义取决于有机体世代的长短(T)与栖境对有机体有利的时间(H)之间的比率(T/H)。这种比值愈小表示栖境稳定。
2.时间上的变异性在一定地点有机体生存的期限内,随着环境条件在时间过程中的便宜,环境的负荷量(K)也随之而变化(也称为“时间上的异质性”)。K的变化可以是周期性的或是可预测性的,也可以是非周期性的或是随机性的。
3.空间上的变异性即栖境是成片的还是分割成不连续的小块。
上述三方面的特性对于种群生态对策的形成均有影响。但其中稳定性常起决定作用。也就是可以把注意力集中在分析(T/H)比值上。但世代的长短/环境有利(T/H)近似于1时,任何一世代的种群对下一代的资源状况无影响。因此,过度拥挤的种群也不会在进化上留下不良的后果。在这种环境下所生存的物种常是积极的进取者,或称为“r类对策”的有机体。相反,在栖境相对稳定的环境中,也就是(T/H)〈1,此时环境负荷量k虽然相对的较为稳定,但是显著的超负荷现象将使k值有所下降。如果当代的种群密度过大时,便会发生不利于以后世代的后果。同时,在这种稳定的栖境中也将会有许多其它的物种迁入并定居下来,造成各种类型的种间竞争,包括捕食现象将因此而激化。在此环境中生活的物种常有高的取食率而在自然选择中被保存下来。这样的生态对策,就称为是k类对策。
三、昆虫的生态对策和防治策略
根据K对策和R对策型的特点,可为害虫的防 治提供参考。对R类对策的害虫其繁殖力较大,大发生频率高,种群恢复能力强,许多种类扩散迁移能力强,常为暴发性害虫,虽有天敌侵袭,但在其大发生之前控制作用比较小。故对此类害虫防治策略应采取以农业防治为基础,化学防治与生物防治并重的综合防治。单纯的化学防治则由于此类昆虫的繁殖能力强、种群易于在短期内迅速恢复,特别是容易产生抗药性,但在大发生的情况下,化学防治可迅速压低种群数量。同时应研究保护利用和释放相应的天敌昆虫,充分发挥生物防治的控制效应。
对K-对策型害虫:虽然其繁殖力低,种群密度一般较低,但常直接为害农作物和林木的花、果实等造成直接的经济损失。故对其防治策略应为农业防治为基础,重视化学防治防治,采用隐蔽性、局部性施药,坚持连年防治,以持续压低种群的密度。
对一些中间型害虫,利用生物防治往往可以收到良好的效果,而不合理地利用化学防治很可能造成再猖獗。
第六节种群数量平衡及其调节理论
影响种群出生率、死亡率和迁移率的各种因素都对种群数量波动起作用。对于昆虫种群波动的重要原因及种群自然调节机制,各学者的观点不同,重要有以下5个学派。
1. 生物因素学派 认为昆虫种群是一个自我管理系统,能按自身的性质和环境状况调节其密度,使种群密度在环境中呈平衡状态,捕食、寄生和种间、种内对共同资源的竞争。即不能决定种群密度的平衡。代表人物(Howard(1911)、Fiske(1911)、Nicholson(1933))。
2.气候因素学派 认为昆虫种群密度的波动主要是当地的气候条件对种群的发育速率和存活率的影响。尤其是对昆虫幼期种群密度影响比较大,所以气候因素是决定因素,而生物因素则影响不大。该学派反对将环境因素分为密度制约因素和非密度制约因素及平衡密度等概念。代表人物(Bodenheimer 1928)
3.综合因素学派认为单纯从生物因素或气候因素来分析昆虫种群数量的波动是片面的,因为他们在不同条件下,不同昆虫种间所起的作用不同。代表Milne(1957)、Buktpob(1955)。
4.动态平衡学派 认为昆虫种群数量能维持在一定水平,是由于种群与生境之间建立了一种动态平衡关系,动态平衡是种群在长期波动中的表现三个过程:代表(Richards(1968)、Clarks(1954))调节过程,当种群密度达到平衡值以上时,个体数的增加受到抑制,当种群密度在平衡值以下时,则存有促进个体数增加的反馈机制。
变动过程,使种群密度偏离平衡值诸因素的作用过程。主要是由非密度制约因素和逆密度制约因素所引起的过程。
条件过程,规定种群平衡密度上下限或引起种群平衡水平变动的诸因素起作用的过程。
5.自动调节学派该学派以内源性因素分析说明昆虫种群数量的波动,认为昆虫的行为、生理、遗传影响种群的的出生率、死亡率和迁移率,引起种群数量的波动。
代表:Ford(1931)、Chitty(1955)、Wedwards(1962)
昆虫种群数量波动的原因十分复杂,在不同的时空条件下,环境因素和自我调节的作用过程都不一样,即引起种群数量波动的主导因素也不相同,应因虫、因地、因时、因环境条件进行具体分析。
第四章 群落生态学
第一节 生物群落概述
一、生物群落的概念:
是指在一定地段或一定生境内各种生物种群构成的结构单元。在自然景观中,不论是在原始的还是在经人为改造的生境内,都有多种植物、动物和微生物生活在一起,它们之间有着复杂的相互联系、相互依存、相互制约的关系。
二、生物群落的基本特征
生物群落的基本特征主要表现在:
①一个生物群落中所有的生物彼此之间通过能量和物质的联系,构成一个复杂的生物种的集合体。
②生物群落与其环境有着不可分割性。
③生物群落中的各个生物成员在群落中的重要性不同。
④同一地域的同一群落都具有其时间和空间的结构的特点。
⑤生物群落的松散性和模糊性。
⑥群落的演替(succession)。群落随时间而变化的动态特征,称为群落演替。
从生物群落整体水平看,群落的基本特征如下:
⑴群落中物种的丰富度(richness);亦即群落中有多少个物种,这是研究群落首先应当了解的问题。(s)
⑵群落中物种的均匀度(evenness);亦即群落中各物种个体数量的分布情况(e)
⑶群落中物种的多样性(diversity of community);物种数、个体数及其均匀度
⑷群落的生长形式及时空结构;
⑸群落的优势种;在一个群落中所有物种的种群起的作用有都是不完全相同的。在群落的发育或能量转换过程中,常由于一个或几个种群的数量、大小和在食物链中的地位深刻地影响着自然环境和群落的组成,似乎能决定整个群落的性质,这样的物种称为群落中的优势种。
⑹群落的营养结构,如食物网结构等。
第二节 群落的结构
生物群落的营养结构概念----群落的格局:生物在环境中分布及其与周围环境之间的相互关系所形成的结构。
一、垂直分层结构
垂直分层格局(stratification pattern),或称垂直结构,即群落的垂直分层现象。大多数群落都有垂直分化或分层现象,即离地面不同高度或水域中不同深度,分布着不同的物种。
Whittaker(1952)年在美国烟雾山不同高度的山坡上作了昆虫群落结构的分析,有七种昆虫分布在不同的垂直高度上,每一物种只局限在一定的高度范围内。对某一物种来说,其中有一个高度上数量最多,然后向上、向下逐渐递减,直到不能生活为止,因此每一个物种的分布都成“钟形”分布曲线
二、水平带状结构
带状格局(zonation pattern),或称水平结构,即群落水平的离散结构。形成局部地区相对高密度集团的小片状分布大致有三个原因:
① 亲代的分布习性
② 环境的差异,因土壤、温度、湿度、植被等环境因子的分布在空间上是不均匀的,昆虫种群就随着点片的生境布局而有了相应的分布型。对于一个群落结构来说,又随着各物种的生物学适应范围的差异,形成了各物种复杂交错的小片状分布。
③ 种间相互关系,群落中物种间的相互关系或结合程度经常不相同,有许多物种经常趋于在一起出现,呈正协调状态,而另一些物种,由于竞争排斥或对环境、资源要求明显差异而相互排斥,呈负协调状态。
三、时间结构
活动性格局(activity pattern),或称时间结构(或格局),即群落的周期性结构,如昼夜格局和季节格局。群落中物种在时间结构上的分化,群落中的物种,除了在空间上有结构的分化外,由于许多自然环境因素如光照、温度、湿度等都有着极强的时间节律,因而物种在时间结构上也有分化。这是物种在长期自然选择过程中,在其生理和生态习性上与周围环境相适应而逐渐建立起来的周期节律。
四、营养结构
食物网络格局(food-web pattern),即群落的营养结构(trophic structure),营养结构可用食物链、食物网和生态锥体来表征。
群落的营养结构是群落内各生物成员之间的最重要的联系,是群落赖以生存的基础,也是了解生态系统中能量流动的核心。分析群落的营养关系,就可以了解其营养结构。营养结构可用食物链、食物网和生态锥体来表征。
⑴ 食物链和食物网
1.食物链:表示植食性动物取食植物、肉食性动物取食植食性动物、另一种肉食性动物又取食该种肉食性动物之间的食物联系。
2.在自然界群落中有许多食物链,这些食物链又共同构成了食物网
⑵ 生态金字塔
生态金字塔又称为生态锥体。群落内各条食物链上的各级(各环)生物的数量、生物量或单位面积、单位时间内所吸收的能量可以反映出群落的营养结构。如果将第一级生物(生产者,多为绿色植物)作为底层,植食性动物和肉食性动物依次向上排列,则呈塔形(或锥形),该种图象称为生态金字塔或生态锥体。
生态金字塔有以下3种基本类型。1.数量金字塔 2.生物量金字塔 3.能量金字塔
A数量金字塔数量金字塔 数量金字塔以各相继营养级别的生物个体数量表示。第一级生物(生产者)为塔底,其数量最多,第二级植食性动物和以后各级肉食性动物数量依次减少,最末级(塔顶)数量最少呈一正的数量金字塔形。但如第一级生物个体很大、数量少(如大树、各种农作物)、而第二级生物(如害虫)和以后各级生物(如寄生性天敌)的个体依次渐小、数量渐多,这样就形成一倒的数量金字塔形。数量金字塔不能表明能量的传递,而且各级生物间数量的变化和差异很大,它过分强调了小型生物个体的数量,故用于表示整个群落的营养结构尚有一定的局限性。
B生物量金字塔•生物量金字塔 生物量金字塔以各级生物的干重来表示各营养级别的生物总量。一般第一级绿色植物的生物量大于植食性动物的生物量,而植食性动物的生物量又大于生物群落中,依此类推,故生物量金字塔也呈一正金字塔形。但在湖泊和海洋的生物群落中,如第一级为十分微小的藻类,其生活周期短,繁殖快,只能积累很少量的有机物质,相反浮游动物取食强度大,体形亦较大,故就现存量来讲,则呈倒的生物量金字塔形,生物量金字塔过分强调了大型生物体的重要性。
C能量金字塔能量金字塔 能量金字塔以各营养级上通过的能流或生产力来表示。构成能量金字塔的依据是单位面积、单位时间所产生的生物量,或作为食物的物质流经食物链的速度。虽然有些生物个体很小,但它们同化和传递的总能量却多于那些比它们大得多的生物,即不受生物个体大小的影响。每个营养级别流出的能量总量要小于流入这个营养级别的能量,因此能量金字塔都呈正金字塔形。能量金字塔提供了群落营养结构的最好的图象。
第三节 群落特性的分析
一、群落的丰富度
群落的丰富度(richness)是表征群落中包含多少个物种的量度,是表征一个群落特性的最基本的一个量度。它常以群落中总共包含拖把种数S来表征。丰富度S愈大,则物种数愈多,生物物种间的关系愈复杂。
一个群落是由许多物种组成的,群落的性质除取决于各成员质的特征也取决于量的特征,这些种群在量方面的特征又分为物种数目和各种群中个体数量两个方面。任何一个群落,僻如对一个孤立的海岛上的群落来说,决定群落丰富度的因素经常有五个:
①历史因素,每个物种移居到这个海岛上来的时间长短。
② 潜在的移居者的数量。也就是移居者原居住地所拥有的潜在能移居的物种有多少。
③ 离潜在移居物种原居住地的距离。
④能容纳这些物种的生境大小。
⑤ 这些物种间的相互关系。如某些种是受抑制正趋于绝灭,还是正在向优势种的方向发展。
在考虑物种数量的同时,还得考虑每个物种的个体数量问题。对这两个方面,自然资源的质与量是一个极为重要的影响因素,这牵涉到资源对这些物种的负荷量问题。如果资源的质对某一二个物种有利,而且数量很大,则群落中的优势种必定突出,这样就使群落的物种结构趋于简单,物种数量少而群落多样性下降。资源质的幅度影响了它所负荷的物种的多少,而资源量的幅度则影响着每一个物种种群的大小,即个体的多少。这里所指资源的质是:如采蜜昆虫对花蜜蜜腺的浓度、蛀木昆虫对树皮的厚薄程度、食谷类仓虫对谷物的干湿、整体可利用的程度等等。
既然自然资源的质与量在一定程度上是群落中限制物种数目和物种个体数量的一个重要因素,那么在一定的自然环境中,群落中所有物种内的个体数目是否存在着一定相对的分布频率呢?从我们日常的田间群落结构的调查工作中,可以发现在任何一个群落中,非优势种虽然种群个体数量很少,但这些物种的数量却是很多的,而优势种虽然种群个体数量很多,但这些物种却并不多。以捕虫灯中所捕的昆虫组成来分析,每一物种内的个体数目与物种数目的频率关系是一抛物线。说明在捕虫灯中绝大多数的物种往往只能捕到少数个体,而个体数量多的物种只有很少几个。从北京来说,在六月底的时候,捕虫灯中铜绿金龟子的数量常是一夜间上千地出现,但这样的物种毕竟是很少的。在麦田昆虫群落结构中,麦蚜可以捕到成千上万,但像这样的种类也只是一至两种。
二、群落的优势度
在整个群落的不同营养阶层中,在数量或功能上占优势、对整个群落的外貌、性质及功能起着决定性影响的物种,称为优势种(dominant species),其余的物种则称为从属种。在评价优势种时,常用Berger和Parker(1974)提出的优势度指数(dominace index)表示,其公式为:
d=Ni/N
式中:d为优势度指数,Ni为优势种的种群数量,N为群落中全部物种种群数量。一般这一指数用对同一营养层中相似的种种类较有参考价值。
三、群落多样性
群落多样性(Diversity of community)是群落中物种数和各物种个体数构成群落结构特征的一种表示方法。
1. 群落特征的表示方法
常用的有种群的密度(Density)、频度(frequeney)、丰富度(richness)、种的相对多度(relative)、多样性指数(deversity index)以及均匀度(evenness)。
A.密度和相对密度(Density and relative density):
密度=样地内个体数/样地面积 D=T/S
相对密度=某个种的个体数/所有种的个体数*100Rd=Ti/Tt×100
B.频度和相对频度
频度(F)=某个种出现的样方数/全部样方数*100
相对频度=某个种的频度/所有种的总频度值*100 Di=Fi/∑F*100
C.丰富度是指群落中出现的物种总数N
D.多度:几种常用的多度等级,不同的学者有不同的分法,但大同小异dominant优势(D)、abundant 丰盛(A)、frequent 常见(F)、Occasional 偶见(O)、rare 稀少(R)、 very rare 罕见(Vr)
2. 群落的多样性指数:
① Shannon-Weiner(香农-维纳)指数(1949)H’= -C∑PilnPi
通常置C=1,这样,Shannon的信息多样性指数就表示为:H’= -∑PilogPi
② S物种数
③ 均匀度(J,)测定
J,=H’/H’max
即在给定物种数S下的完全均匀群落的多样性指数最大多样性
S为群落中的种数
④Hurlburt (1971) 种间相遇机率(PIE)
PIE=∑(ni/n)[(N-ni)/(N-1)]
⑤Simpson (辛普森多样性指数)的多样性量度
可以用D=1-∑Ni(Ni-1)/N(N-1)作为集合多样性的一个量度.当集合只是一个随机样本,而且它来自具有同样多种的某个无限大的母体时D可用下式计算即 D=1-∑Pi2D为辛普森多样性指数,S为物种数,Ni为群落中第i种物种的个体数,N为群落中物种的个体数,i为1、2、3、、、、S。
四、群落的稳定性
群落的稳定性(stauility of community)是指群落抑制物种种群波动和从扰动中恢复平稳状态的能力。它包括群落现状稳定性、时间过程稳定性、抗扰动能力稳定性和扰动后恢复平稳的稳定性等4种情况。群落稳定性和生态系统的稳定性具有同一概念,主要包括两种能力,即抵抗力和恢复力。所谓抵抗力即抗变能力,表示群落抵抗扰动、维持群落结构和功能、保持现状的能力。所谓恢复力表示群落在遭受扰动以后恢复原状的能力。恢复得越快,群落也越稳定。
第四节 群落的演替
一、群落演替的基本概念
群落演替(community succession)又称生态演替(ecological succession),是指在一定区域内,群落随时间而发生变化,由一种类型转变为另一种类型的生态过程。或者说是群落经过一定的发展历史时期,由于物理环境条件的改变,从一群落类型转变成另一类型的顺序过程。
群落演替包括3个方面的基本观点:
① 演替是群落组成有规律的、顺序的向一定方向发展的过程是可以预见的。
② 演替是由群落引起物理环境改变的结果,即演替是由群落控制的,一般是不可逆的。
③ 演替以形成稳定的生态系统,即由顶极群落所形成的系统为其发展的顶点。
在特定的地区内,群落由一种类型转变为另一种类型的整个取代顺序,称为演替系列(sere).演替过程可分为若干不同阶段,称为系列更替(serechange)。在演替初期称为先锋期(pioneer stage);演替中期称为发展期(developmental stage);发展到最后的稳定系统,称为系列顶极(seral climax)或顶极群落(climax community)。
演替是群落中的有机体和环境反复相互作用,发生在时间、空间上不可逆变化的发展过程。当第一个先驱的有强适应力的物种侵入环境后,使土壤、小气候等条件改变,就为第二个物种和第三个物种的进入创造了条件。第二、三个物种在继续改变环境条件的同时,就可能抑制,甚至排挤那个先驱的物种。所以群落的演替不但是由于构成群落的物种的内因,而且还更多地涉及环境外因。变化过程中,环境梯度的变化与种群间梯度的变化交织在一起而形成了群落特征梯度的变化。从这个意义上说,演替就是时间上的生态群落的梯度。
生态群落梯度的变化
每一个生态群落梯度的变化是有一定规律的。例如在棉田的群落中,随着棉株不同发育期的生理生态变化,群落中的昆虫物种的组成,也发生了相应的季节改变。在棉花的出苗期主要以地老虎、蛴螬、蝼蛄、金针虫等地下害虫为主;苗期主要以刺吸式口器的棉蚜、棉红蜘蛛、蓟马、盲蝽为主;蕾铃期则改变为食叶蛀蕾的棉铃虫、红铃虫、棉大卷叶螟、造桥虫等害虫,在这些不同的害虫发生时期中,同时出现了与此相联系的天敌种类,并共同组成不同棉花生育期的昆虫群落。掌握棉花生长过程中生物群落的结构与季节性演替,有助于在不同阶段制订有效的增产与防治措施。
二、群落演替的类别
1、根据演替出现的起始点,可以将群落演替分为原生演替(primary succession)和次生演替(secondary succission)两类。原生演替开始于从未被生物占据过的区域,又称初级演替。次生演替指在原来就有生物群落或曾经被生物占据过的地方发生演替,又称次级演替。
2、 根据引起演替的原因,可以将群落演替分为内因性演替(endogenetic succession)和外因性演替(exogenetic succession)。前者是指由于群落内部不同物种的竞争、抑制或生命活动,而改变环境条件引起的演替。后者是指非生物因素变动引起的演替。
3、根据群落代谢的特征,可以将群落分为自养性演替(autotrophic succession)和异养性演替(heterotrophic succession)。前者是指群落中主要生物以增加光和作用产物的方式进行演替,如由裸岩→地衣→苔藓→草本植物→灌木→森林的过程。后者则相反,如受污染的水体,在那里细菌和真菌的分解作用特别强,使得群落中有机物的量由于腐败分解而逐渐减少。
三、群落演替的过程
1、群落的演替过程简单地说可以分三个阶段
⑴侵入定居阶段:首先是一些强有力的先驱物种侵入定居并获得成功,刚开始时可能只有这些物种中的少数个体能幸存下来并繁殖后代,或只是在很小的一些生境中存活下来,但这些初步建立起来的种群却对以后环境的改造,和以后相继侵入定居的同种或异种个体起了极为重要的奠基作用。
⑵竞争平衡阶段:群落在发展,种群数量在增加,当有了一定数量的物种进入这个生境以后,生活小区逐渐得到改造,资源的利用逐渐由不完善发展到尽可能的利用,种内与种间的竞争在激烈的斗争中渐渐趋向于平衡。
⑶顶极平衡阶段:在竞争平衡阶段中,物种间的关系在变化,群落在发展和演替,一直达到“顶极”阶段。这时候,优势种的特征已相对地稳定下来,使整个群落环境之间保持着一种动态平衡。
2、在群落演替中物种组成的相互关系方面,Wilson(1969)提出了四个阶段。
⑴互不干扰阶段 群落中物种从无到有的最初阶段,这时候物种数目和各种群密度都很低,还不可能在自然资源的利用上有什么竞争,对捕食性或寄生性天敌来说,还不存在开发利用这些寄主的问题。
⑵相互干扰阶段 这是一个由于竞争而相互平衡的阶段,在竞争中有不少物种侵入并定居下来,且得到了繁盛的机会,而另一些物种则被排挤而趋于灭绝。
⑶共摊阶段在这阶段中那些能很好地利用自然资源而又能在物种的相互关系中共存下来的物种就得到了发展。它们从不同的角度利用自然资源,共摊自然资源。
⑷进化阶段物种中的协同进化使自然资源的利用更为有效,有时可能再增加一些共存的物种使群落在结构上更趋紧密。
四、群落演替的特征
1、演替的方向性 大多数群落的演替都有着共同的趋向,而且是不可逆的。演替的方向一般是从低等生物逐渐发展到高等生物;从小型生物发展到大型生物;生活史从短到长,群落层次从少到多;营养阶层从低到高,由简单到复杂,物种从少到多,种间关系从不平衡到平衡,从不稳定趋向稳定
2、演替的速度:一个先驱物种中的某些个体,要在一个荒原上形成种群,再从它的基础上发展成为一个初级的群落,这可能是一个艰难的长期自然选择过程。一般来说,这个过程是极为缓慢的,当一个初级群落建立起来以后,每一个定居下来的新物种都面临着繁殖、扩散、巩固的问题,物种之间开始了激烈的竞争,也就是对有限的资源,或生境空间展开争夺。在严酷的生存斗争中,群落的物种组成是不稳定的,经常在数年中或数十年中就会更换一系列的物种。但稳定平衡还是演替的必然结果,在交错复杂的种间竞争与对自然环境的斗争中,毕竟会是一些强有力的优势种获得主导地位,因而使演替的速度缓慢下来,最后以群落的稳定平衡中只存在某种相对的波动。
3、
三. 群落演替的效应
四.
演替的效应:有不少生态学家认为群落中的物种在自身的发展过程中,经常对生境产生一些不利于自身生存而有利于其他物种生存的因素,因而在演替中创造了物种替代的环境条件。
很多种植食性害虫,由于为害作物经常破坏了它自身生活的适宜生境,毁灭了作为其生境和食物的寄主植物。但同时为另外一些处于相继的营养阶层的物种造成了繁盛的条件,导致了群落演替过程中物种组成成分的更替。
第五节 生态位
一、生境、生活小区和生态位概念
生境(habitat):生物的居住场所,即生物生活的地方,或称为栖息地。
生活小区(biotype):在生境内主要环境状况一致的。所谓生活小区是指具有相似土壤和气候条件,栖息着一定动植物群体的场所。生活小区是在生境内生物群落的居住单位。有时,就同一物种而言,可有不同的生活小区,如棉铃虫可以分别居住于棉田、麦田、玉米田等。
生态位(ecology niche):是由美国学者约翰森提出来的,用以描述不同物种在同一生活小区内可以占据资源的部位。一般强调生物有机体在群落中的功能作用。
一个动物的生态位表明它在生物环境中的地位及它与食物、天敌的关系。并与种间竞争的概念相联系。
二、生态位的分类:
按影响生态位的环境因素个数,可分为一维生态位(one-dimensional niche)、二维生态位(two-dimensional niche)、三维生态位(three-dimensional niche)和多维生态位(multidimensional niche)。
一维生态位常用生态位宽度(niche breadth)和生态位重叠(niche overlap)两项定量指标,比较群落中各物种占据空间的大小和利用资源的多少。如某物种对温度的适应幅度广,或在寄主植物上分布范围大(即占据资源范围大),则该物种的生态位宽度指数大,反之则小。如某物种与另一物种适应于同一温度范围,或分布于寄主植物的同一部位(即利用相同等级资源)的数量多,则这两个物种的生态位重叠指数大,反之则小。
1、
生态位宽度:
美国学者莱文Levins(1968)根据物种对环境幅度的反应,首先提出。用以表明一个物种在具有不同数量单位的资源序列上的分布。
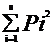
B=1/S*
B为生态位宽度指数
S为资源系列的等级数
Pi为某物种在第I等级资源数量占总资源数量的比值。
B值的变化范围为1/S〈 B〈 1 Lij和Lji分别为第I物种重叠第j物种和第j物种重叠第i物种的生态位重叠指数;Bi和Bj分别为第I物种和第j物种的生态位宽度指数。
PihPjh分别为第I物种和第j物种在h资源等级中利用资源占的比例。S资源系列等级数。
2、
生态位重叠
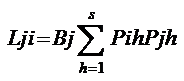 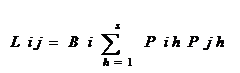 基于Levins生态位宽度的重叠指数(L)其公式 基于Levins生态位宽度的重叠指数(L)其公式
Hurlbert(1978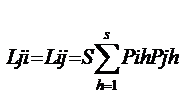 ): ):
Lij和Lji分别为第I物种重叠第j物种和第j物种重叠第i物种的生态位重叠指数;Bi和Bj分别为第I物种和第j物种的生态位宽度指数。PihPjh分别为第I物种和第j物种在h资源等级中利用资源占的比例。S资源系列等级数。
第五章生态系统生态学
第一节 生态系统的基本概念及农业生态系统的特点
一、生态系统的概念
生态系统(ecosystem)指在一定空间内,生物群落与非生物环境之间通过物质循环、能量流动和信息联系等过程,结合而成的生态单元,即生物群落与其环境条件所形成的统一整体。
生态系统不是静止不变的,而是一个动态的系统。如果系统中某一要素(生物群落中的某些成员、非生物环境中的某些生态因素)发生变化时,就可能通过物流、能流、信息流等相应改变,而影响其它因素,进而引起整个生态系统的变化。
二、
1、生态系统的基本结构
生态系统结构是指系统中各种成分及其相互关系和联结的形式。
从组织水平考虑,生态系统的生物成分可以分为个体、种群、群落和生态系统。
从营养功能考虑,生态系统的结构包含以下四个主要成分。
1、非生物环境:包括土壤、水等的理化性质和成分,构成植物生长和动物活动的空间;物质代谢的环境,生物代谢的媒介物质;气候条件。
2、生产者:(productors)即自养生(autotrophs),是指能利用太阳能,经过光和作用,将CO2和水合成碳水化合物的绿色植物。它可通过进一步合成脂肪和蛋白质供其自身的建造,也为生态系统中的消费者和分解者提供营养。
3、消费者:(consumers)指那些不能从无机物质制造有机物质、直接或间接依赖生产者制造的有机物质生活的生物,即异养生物(heterotrpths)。
4、分解者:(decomposers)指小型消费者(microconsumers),又称腐食者(saprothroths),主要是包括细菌和真菌等小型异养生物。
三、
生态系统的基本功能
生态系统的基本功能是指系统之中和系统之间的物流和能流的运动过程。与生产者、消费者和分解者3种成分相对应,其基本功能分为生产、消费和分解3个过程。由于消费过程是食草动物和食肉动物将生产者已经生产合成的有机物质及储存于其中的能量,按照一定途径在生态系统中运动,所以生态系统中实际上最基本的功能过程是生产过程和分解过程。
四、生态系统的能流和物流
能流(energy flow):
指在生态系统中各营养级生物间能量的输入、转换、传导、消失的过程,它是一个单向流动的过程。Lindema(1941)提出了能流的概念和“1/10定律”,即从上一营养级到下一营养级的能量转化率是10%。
物流(material flow):指物质在生态系统中流动的过程。
关系:能流和物流过程是相互联系的,储存于化学键中的能量,在物质流动和变化的过程中,总是伴随有能量的流动和变化,但二者又有所区别,,能量只能使用一次,故能流是单向的,而物质可以反复被利用,谷物流是可以循环的。生态系统的基本功能是物质循环(营养循环)和能量流动,如果为管理措施不当,其基本功能遭到破坏,生态系统将失去平衡。
五、生态系统的类型
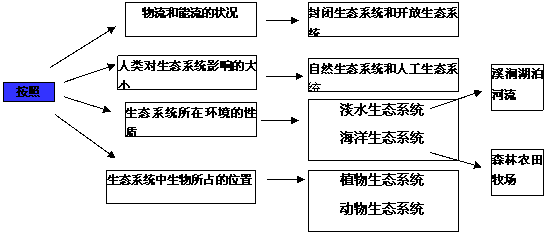
六、农业生态系统:
农业生态系统(agroecosystem)指人类从事农业生产活动,所形成的生态系统。广义的农业生态系统包括种植业、饲养业和农畜产品加工业等活动所形成的系统。狭义的农业生态系统则是指由于各种种植业所形成的农田生态系统。
农业生态系统与自然生态系统相比较:主要的不同。
1、种植的植物单一,其生长发育进度整齐,植物种间的竞争较少,各农田生态系统的分界明显。
2、以人们选定的种植植物为中心所形成的生物群落中,生物类群贫乏,营养层次简短,食物链环数目少,系统内部自我调节的机能较差,系统的稳定性差,易受不良环境因素的影响。
3、由于种植的植物(生产者)种类单一,数量多,往往导致一些一级消费者(如害虫)数量多,危害重,成为优势种害虫,而需要防治,尤其使用化学防治法,常使这一系统遭到破坏,失去平衡。
4、种植的植物生长时间短,如一年生的作物,蔬菜和多年生的果树,而使群落的演替时间短或是不连续。
5、由于系统内的一些产品(如种子、果实、叶、根等)被人们收获而离开系统,故必须采取施肥、灌水等手段。以维持系统中正常的物流和能流,保证高的生产力。
6、生态系统的机构因社会(人类)的需要、经济效益而发生变化,故实际上农业生态系统是社会—经济—自然生态系统组成的复合系统。所以人们对农业生态系统的影响,可能是积极的建设作用,也可能是消极的破坏作用。设计农业害虫综合治理的原则:以生态系统管理为理论基础,使生态系统和农业生态系统向着有利于人类的方向发展,以农业防治措施为基础,充分发挥自然控制因素(如作为第二级消费者的害虫天敌、不利于害虫发生的气候因素等)的控害作用,协调运用生物、化学、物理、遗传等防治措施,将害虫的危害控制在经济允许水平之下,并逐渐向着以农田所有有害生物为防治对象的、以利用自然控制因素控害为主的、区域性的综合治理的方向发展。(IPM)
第六章生物多样性及其保护
第一节 生物多样性的结构与分布
一、生物多样性的
一、 定义
生物多样性(biodiversity):是生物及其环境形成的生态复合体以及与此相关的各种生态过程的总和。它包括所有植物、动物和微生物,它们所包含的基因以及由它构成的复杂生态系统和它们形成的过程。
一般来讲,我们从这几个方面来考虑
种内---遗传多样性
种间---物种多样性
共同---生态系统多样性
(一)、遗传多样性
1、遗传多样性定义
广义遗传多样性:地球上的所有生物所携带的遗传信息的总和,即各种生物所拥有的多种多样的遗传信息。
狭义遗传多样性:种内个体或一个群体内不同个体的遗传变异的遗传总和,也称基因多样性。
2、
种群变异度取决于那些因素
A.基因库中基因的数量
B.每个多型基因的等基因数量,多型基因的存在允许种群中个体的基因是杂合的,即从每个亲本中接受基因的不同等位基因。
遗传变异使得物种能够适应变化的环境。一般来讲,珍稀物种比广泛物种有较低的遗传变异,因而在环境变化时更容易灭绝。
(二)、物种多样性
1、物种多样性定义及应用
A、形态学定义:物种是在形态、生理或是生物化学方面有别于其他类群的一类个体的集合。DNA序列已越来越多地用于形态上极相似的物种的分类。
B、生物学定义:物种是一类其内可以繁殖,而与其他类群个体生殖隔离的个体的集合。
应用:形态学定义是分类学家广泛使用的定义,生物学定义则是进化生物学家广泛使用的定义。因其兴趣在于可测量的遗传关系而非物种的某些主观的外部特征。实践中,由于物种生殖资料的匮乏,物种的生物学定义难得使用。
2、物种多样性研究方面还存在那些问题呢?
A、物种多样性
A、 从何而来?
B、 物种多样性怎样保持?
C、 物种多样性怎样测度?
D、 物种多样性生态功能如何(如果存在)?
E、 人类如何依赖物种多样性?
F、 物种多样性今天是怎么变化的?
(三)、生态系统的多样性
生态系统的多样性的
定义:生态系统多样性是指生物圈内生境、生物群落和生态过程的多样化。
生态系统由植物群落、动物群落、微生物群落及其栖息地环境的非生命因子所组成。群落内部、群落之间以及栖息环境之间存在着极其复杂的相互关系。其主要的生态过程包括能量流动、水分流动、养分循环、生物之间的相互关系(竞争、捕食、共生和寄生等)。其中资源的数量和质量、物种之间的相互作用的结果形成了群落和生态系统的复杂性。而系统中的营养多样性对生态系统的功能是不可或缺的。
二、生物多样性的生物地理格局
物种多样性在地表的丰富度因地而异。这是由于地球表面的物理因子的变化影响的结果。物种的数量通常是随着物理变量的变化有所增减,也即地理上的梯度变化。
1、纬度梯度
从极地至赤道,生物多样性明显增加,这是生物地理学中最明显的格局。这个是受到太阳辐射、降水、温度等的季节性波动和其他因子的变化的影响,也称为物理梯度的影响。最大的生物多样性出现在热带森林。另外,生物多样性在地区之间变异极大,一般来讲,热带地区的生物多样性在新热带区最多,在热带非洲地区最少。
2、垂直高度梯度
随着海拔的变化,物理条件、气候条件也随之变化。很多物种的丰富度随着高度的增加而降低。亦有人认为,垂直高度梯度与物种丰富度的纬度梯度相似。
3、热点地区
保护生物学中用“热点地区(hot spots)”来突出生物多样性所面临的威胁的自然状态,并将其锁定在较小的区域。这个概念显示了生物多样性因地域的集中而变的更脆弱。主要应用于热带和亚热带地区。我们昆虫的资料主要是蝶类。
三、物种---面积关系
物种—面积关系或是“岛屿学说”是生态学家长期以来一直感兴趣的关于生物多样性自然变化的一个方面。(岛屿上的物种数随着岛屿面积的增大而增加,且距大陆越远,物种数越少)对物种数随面积和隔离度变化的原因,主要的有平衡假说和栖息地异质性假说。对生物来说,栖息地的片段化和隔离,使破碎的生境类似岛屿,物种在岛屿之间的迁移扩散很少,如湖泊、间断的草甸、保护区和片段化的森林等,称为栖息地岛屿。岛屿上的物种数量是新种迁入定殖和消失(灭绝)之间动态平衡的结果。岛屿上物种迁入率随物种数增加而渐降,消失率却渐增。迁移扩散在决定物种数上起着重要作用。隔离度越大,迁移率变小,平衡物种数也小。(岛屿面积变小,物种消失率增加)。原因是生存面积不足而导致遗传多样性丧失和适应力降低,种群变小,增加了种群随机灭绝的概率。
栖息地异质性假说(Lack,1969,1976)
面积增大包含了更多类型的栖息地,因而有更多不同的物种存在。物种随岛屿面积增加而增加的原因是由于栖息地也增加的结果,而非平衡假说中岛屿的面积效应。
联系:两个学说有其互补性,对美拉尼西亚群岛和波多黎各群岛的蚁类研究(Wilson,1961,Torres和Snelling,1977)表明。美拉尼西亚群岛上的蚂蚁种类数量随岛屿面积的增加而增加,但在波多黎各群岛18年的记录中,迁入量一直超过灭绝量,岛屿面积对蚂蚁种类的多少没有什么影响,栖息地异质性却是蚂蚁种数的最主要的影响因子。
四、 生物多样性的测度
在一个单一群落中,物种的数目通常被描述为α多样性,α多样性与我们讲述的物种的丰富度概念(群落中的物种数,S表示)最为接近,可用以比较不同生态类型栖境中物种的数目。α多样性的测度还有若干定量指数,如Simpson指数、种间相迁率、Shannon-Wiener指数、均匀度程度等。
β多样性用来描述物种组成随环境梯度改变的程度。β多样性的测定值可以指示生境被物种分割的程度,可以用来比较不同地段的生境多样性。β多样性和α多样性一起构成了一定地段的生物异质性。
β多样性的测度方法有多种,我们看其中的一种,即Cody指数:
βc=[g(H)+L(H)]/2
g(H)----随生境梯度H增加的物种数目
L(H)-----随生境梯度H失去的物种数目,即在上一个梯度中存活在而在下一个梯度中没有的物种数。
βc指数通过对新增加和失去的物种数目的比较,使人们能获得十分重要的物种更替概念。
第二节生物多样性的危机
Diamond(1989)总结了物种灭绝的4种原因,称之为“魔鬼四重奏”即生境的破坏和片段化,对动植物的过度掠取,外源种入侵及以上3种原因导致的次生灭绝效应。
所谓用进废退,大家考虑一下,物种的灭绝对我们有什么样的提示??
一个例子:最敏感的昆虫在新的农药诞生以后呢,最容易被杀死,然后留下的是那些生命力强的,于是有了抗性有了“3R”,我们又一次不段的去探索新的方法,从而有了大量的新的投入。
人们在种植一些作物时往往是要留一小块地来种那些能保留敏感昆虫的作物,为什么?
一、未来昆虫的灭绝
岛屿生物学的理论可以用来预测栖息地岛屿上昆虫的命运及栖息地损失,特别是森林砍伐对全球物种丰富度的影响。
用S表示物种数量,A表示岛屿的面积,则有
C和z是常数,C值主要取决于测度单位及栖息地和生物群落,z为无维参数,用以测度随面积增大而出现的物种数的增加。
不看过去的数字,我们从1993-2003年的10年间,大约有25万个物种将会灭绝。而且在随后的50年内将有数十万个物种走向灭绝。
二、生物多样性与气候变化
全球变暖是个无可争议的事实,到2050年,大气中二氧化碳含量将达到工业化前水平的一倍,预计全球气温将升高约1.6摄氏度,海平面将上升约37厘米。----陆地农业将和海洋农业结合(预测)!由于气候变化而影响昆虫种类,不同的种可能以不同的方式对气候变化作出不同程度的反应。这将导致种间平衡被打破,使某些重出现局地灭绝。如当某些当前是无害的种类比它们的寄生性或是捕食性天敌扩散更快时,这些种类将由无害变为害虫,而其天敌有可能灭绝。对气候变化的不同反应还可能形成基于新的种间关系的动植物群落,导致一些共生昆虫(像专性传粉者)的丧失和新病害出现且流行次数增加,从而可能造成局部灭绝。
第三节 生物多样性的保护
生物多样性的保护着重从四个方面入手,即基因、物种、生态系统和景观。
一、生物多样性保护的理论
(一)、岛屿生物学理论
我门依据这种理论提出了若干保护区设置的原则。Diamod等(1976)根据平衡假设,提出了自然保护区设置的几点原则:
① 保护区面积越大越好
② 单个保护区要比面积相同、但分隔成若干的小保护区好
③ 若干个分隔的小保护区越靠近越好,排列越紧凑越好,线形排列最差
④ 有廊道连接若干分隔的小保护区比无廊道连接的好
⑤ 圆形的保护区比条状的保护区好
(二)、异质种群(metapopulation)理论
定义:Levin(1970)提出了异质种群理论,即由经常局部性灭绝,但又重新定居而再生的种群所组成的种群。
区别:异质种群理论主要研究局部种群灭绝,再定居规律及其生存力。岛屿生态学从群落水平上研究物种数的变化规律,而异质种群理论则从种群水平上研究局部种群消失灭绝规律。前者是着重格局(pattern),而后者是强调过程(process),二者既有区别,又有相同之处。物种数目的变化最终还要取决于单个独立物种的灭绝或再定居。故从种群水平上探讨物种灭绝规律对于理解群落的变化是至关重要的。
1、种群的基本模型
Levin从一个全新的角度重新研究了局部种群的灭绝和空生境斑块被重新侵入的问题。
dp/dt=mp(1-p)-ep
P0=1-e/m
P指种群占据的生境斑块与总的生境斑块数的比率。
M表示与所研究物种有关的定居系数
e表示与与所研究物种有关的灭绝系数
上面是异质种群理论最经典的模型。它描述了一个最简单的异质种群随时间变化动态。从本质上讲,这个模型类似于种群生态学中描述单种种群增长的logistic模型,可将上式改写成下面完全等价的形式:
dp/dt=(m-e)p[1-p/(1-e/m)]
这里呢,差值(m-e)是一个充分小的异质种群的增长率[即p(t)]是充分小的],1-e/m是与logistic模型中的环境容量等价的值。如果m>e,则1-e/m必定是p(t)的稳定平衡值,也只有在m>e时,异质种群才能持久生存。
2、异质种群的生存
在自然界中,异质种群的格局分布并不是绝对独立的,而且,局部种群之间的相关性和迁移扩散能力是相关联的。局部种群之间独立性越强,越能较好地对抗环境随机性带来的相关性灭绝的影响,但同时却不利于再定居,这样对于异质种群的生存力又会有不利的作用。(Gilpin,1988)可见,隔离度对确定异质种群的生存力至关重要。过分的隔离会使局部种群之间孤立起来,无法实现扩散和在定居,不利于持久生存;反之,局部种群之间相距太近,关联太强,相关性灭绝的概率就越大,也不利于异质种群的生存。只有适度的隔离才有利于异质种群的长期生存。
3、生物多样性保护原则
异质种群理论在保护区设置上原则上与传统的岛屿生物学理论所倡导的观点有很大的不同,它认为建立若干个小的保护区要比建立与其面积之各相等的一个大保护区为好。实际上呢,由于人类的干扰,栖息地片段化已成为事实,在世界各地,建立一个大的完整的保护区很困难,所以呢,在这种情况下,异质种群理论在生物多样性保护上更具有现实意义。
(三)、最小生存种群理论
当种群过度碎裂和隔离后,每个格局种群的个体数量变的很小,且与其它局部局部种群孤立开来,这样,它的灭绝将是永久性的,即无法达到其他局部种群个体的在定居。当所有其他局部种群相继灭绝,只剩下最后一个局部种群时,该物种便难逃灭绝的厄运,一些濒危物种就是这样的例子。
最小生存种群理论原因
种群一旦变的太小,原来大种群动态研究中忽略的随机因素便上升为决定因素,使之濒于灭绝的危险中。这可归纳为三个原因:
1、缺乏遗传变异性、近亲繁殖和遗传漂变导致的遗传问题。
2、出生率和死亡率的随意性导致种群数量的不稳定。
3、捕食、竞争、疾病和食物供应的变化导致的环境波动,还有不规律发生的单一事件。如火灾、洪水和干旱等等。
一个小的种群,它究竟能活多久?Shaffer(1981)将保证一个种群存活所必需的个体数量定义为该物种的最小生存种群(minimum viable population,MVP):最小生存种群是任何生境中的任一物种的隔离种群,即使是在可预见的种群数量、环境、遗传变异和自然灾害等因素的影响之下,都有99%的可能性存活1000年,换而言之,最小生存种群是在可预见的将来,具有很高生存机会的最小种群。
Franklin(1980)认为50个个体应是最低限度,即动物群体可在每代丧失2%~3%的遗传变异性的情况下维持生存。按照Wright(1931)提出的每一世代(F)成体数量(Ne)杂合性降低的公式 F=1/(2Ne)。一个具有50个个体的种群,每代仅会丧失1%的变异性。所以得出以下结论:需要多少个体才能保持种群的遗传变异呢?
50/500法则:隔离种群至少需要50个个体,为保持遗传变异性最好拥有500个个体。
二、昆虫的保护
(一)、保护濒危物种有两种不同的策略:
原地保护:即在该物种的自然生境或亚自然生境中施行保护
异地保护:如动物园、植物园,但只有极少数几种昆虫采取这样的措施。
(二)、在昆虫的自然保护中,我们必须注意以下问题:
1、所有对昆虫重要的栖境都要保留。
2、栖息地的面积应足够大而且保持一定形状。
3、应保持足够数量的生境类型、物种的个体各不同栖息地之间的迁移以确保其长期生存。
4、采用适宜的栖息地管理策略。
(三)、栖境的保护
昆虫的保护中最紧迫和最重要的是热带雨林的昆虫的保护,因为这些森林虽然覆盖了地球陆地表面的8%,但却含有全世界物种数的50%~80%,但是热带雨林的年砍伐率大约为1%所以:必须保护,最好的人工保护栖境方法是建立自然保护区,或是自然公园等。
第四节生物多样性的保护和可持续性发展可持续性发展
一、生物多样性保护的环境经济问题
什么因素使得人类以一种破坏性的方式来行动?经济因素,一般来说,自由交换的成本和收益是由交易双方承担和接受的。但在某些情况下,不直接参与交换个体也会付出代价或获取利益。在这些外在的成本或是收益称为经济活动的外部后果。最明显和最容易被忽视的一种外部后果就是环境的损坏。
成本和收益的分离正是市场失败的核心问题:一种经济活动的成本由民众担负了,而利益却集中到少数人身上。这是经济和生态冲突的必然原因。许多的自然资源,如清新的空气、清净的水、肥沃的土地、稀有的物种、甚至美丽的风景都是真个社会拥有的公共资源,而这些资源往往不被赋予金钱价值。公众、企业、政府使用或损坏这些资源时并不计入企业的内部成本而付出极少,有时甚至无须付出,对生物资源的利用也是如此。未经过市场流通而直接消费,因此呢,我们寻求一种评价方法,来计算生物多样性对国民经济的贡献。在众多的方法中,主要看McNedy(1988,1990)提出的方法。在这种方法的框架内部,价值被分为直接价值和间接价值。直接价值是那些由人类直接收获或是使用的产品,间接价值指生物多样性所提供的利益,不涉及资源的收获或是毁损。
二、生物多样性的直接价值
直接价值进一步分为消耗使用价值和生产使用价值:就地使用的物品体现着消耗使用价值,进入市场的产品体现着生产使用价值。
消耗使用价值:生物多样性为人类提供了基本食物。另外一项------药物也是人类生存的关键,而大多数的药物是由动植物和微生物提取或是合成的。
生产使用价值:生物多样性还为人类提供了多样的工业原料。如木材、橡胶、油脂等等。
目前,木材是最重要的产品之一,其生产值超过750亿美元(Reid&Miller 1989)。此外,许多物种的最大生产使用价值在于它可以作为工、农业以及农作物遗传改良的原材料。野生物种也可用作生物防治的资源。通过寻找有害作物的天敌来防治它的扩散。最后,自然界做为药物的重要来源也产生了无法估计价值。
三、 物多样性的间接价值
间接价值指生物多样性的另外一些与生态系统功能有关的,如环境作用(environment process)和生态系统维护(ecosystem services)。这些过程和维护能确保自然产品的持续生产,但在使用过程中却不受损坏。
生物多样性的间接价值也可以看做环境资源的价值,其意义可归纳为以下2个方面。
(一)、非消费使用价值
生物群落提供了多种在使用中不被消耗掉的环境服务,如野生昆虫给农作物传粉的价值,这种使用价值算起来相对容易些,可由其传粉活动所增加的农作物产值来估算。但确定生态系统的其他使用价值要更困难些,特别是在全球尺度上。
主要集中在七个方面:
1、生态系统生产力
在植物和藻类的光和作用这一能量的不断转化中,其使用价值是很难估算的,但我们破坏了一个地区植被以后呢,就损坏了生态系统转化太阳能的效率,最终导致植物生产力的丧失和生活于该地区动物群落的萎缩。目前我们在生态系统多样性研究中一个重要的问题就是生物群落中个别物种的丧失,会如何影响生态系统的功能和作用。
2、保护水土资源
生物群落在保护流域、缓冲旱涝对生态系统的冲击和保持水质等方面至关重要。尤其是植被的多少可以对流区水土的保持起决定性作用。在集水区内发育良好的植被具有调节径流的作用。相反,砍伐、垦荒和其他人类活动减少植被时,水土流失,洪水泛滥等等现象就会严重。
3、气候调节
植物群落在调节局部区域、地区以及全球气候方面也起到恶劣重要作用。在局部区域层次上,树木提供遮荫处,蒸发水分,从而在夏季能降低气温。树木做为风障,还能大大减少冬季建筑物的热能损失。在地区层次上,植物蒸腾作用使水循环到大气中,再以雨的形式返回地面。
4、污染物的吸收和分解
生物群落能降解和固定污染物,但是当生态系统被破坏后,我们所得的要用来处理我们所遗留的,也许还远远不够,有人说21世纪最糟糕的发明是—一次性塑料袋。
5、种间关系
许多物种因有生产使用价值而被人类利用。然而,他们的持续生存却依赖于其他野生物种。因此,一个对人类无多少直接价值的野生物种的减少,可能导致具有重要经济意义物种的相应减少。如农田节肢动物的群落研究表明,那些对人类既无害又无益的中性昆虫(如水稻田内主要是指一类如蚊、蝇、弹尾虫等),对天敌种群的发展和维持,尤其对早期天敌种群的建立有着不可或缺的作用。(吴进才 1995)
6、休闲和生态旅游
这里主要是指如远足、摄影、观鸟等活动来欣赏大自然。而生态多样性是这二者的基本条件。
7、环境监测
8、对化学毒物特别敏感的物种能作为检测环境健康的“早期预警系统”。如苔藓生长在岩石上,吸收酸雨和空气中污染物的化学物质,是最显著的指示物种。
(二)、备择价值
物种的择价值是指物种在未来某个时候能为人类提供经济利益的潜能。当社会需求发生变化时,满足这些需求的方法也必须改变,解决问题的途径往往是寻找新的自然生物:昆虫学家寻找能用于生物防治的天敌昆虫,微生物学家寻找能促进生化过程的细菌,动物学家呢,寻找能更有效的生产动物蛋白、对环境损害较小的物种,药物学家则致力于收集和筛选野生物种中有药用价值的成分。如果生物多样性减少了,发现和利用新物种的能力也随之减少。研究工作者的最大任务就是寻找一种现存物种的潜在利用价值。
第五节 生态农业
一、农业生态的定义
定义:生态农业是运用生态系统中的生物共生和物质循环再生原理,采用系统工程的方法,吸收现代科学成就,因地制宜,合理组织农、林、牧、副、渔生产,以实现生态效益、经济效益和社会效益协调发展的农业生产体系。简单得讲,生态农业就是以生态学原理为指导,建设一种既有利于资源和环境的保护,有能促进农业生产发展的农业生产体系。
二、现代农业的负效应:
有人将现代农业称为“工业化农业”或“石油农业”。现代农业高投入、高产出的生产方式使土地生产力和农业生产力得到了飞速的增长,对解决人类食物供应的贡献上巨大的,但由于在资源、环境、生产成本等方面造成的负效应,其弊端日益明显:主要在以下5个方面
1、能源过度消耗:在美国,每生产1000J能量的水果与蔬菜需要投入2000J的石油能,生产1000J能量的动物蛋白需要投入20~30KJ的石油能,如果按这个标准来估算,全世界都采用美国的生产方式与营养水平,每年将消耗60000亿L以上的石油,而目前探明的石油储量只够全世界人口使用十几年。----西气东输
2、水资源消耗----南水北调
3、生产成本增长:一个简单的数据,1950~1985年的35年间,世界化肥用量增加恶劣8.29倍,总耗能增加了5.9倍,而此间谷物仅仅增加了1.68倍。
4、污染加剧----我们看看自己周围!
3、 其它负效应,如大规模的专业化和规模化生产的发展,使农作物种植趋向于单一化连作,使生态系统中生物多样性遭到消弱,其不稳定性和脆弱性加大,对自然灾害及病虫害抵御能力下降。对土壤高强度的垦殖及用养失调造成的水土流失、土壤退化与沙化、草原退化等,已成为现代农业不可忽视的严重问题。
三、生态农业的基本原理:
中国生态农业是从总体上充分发挥地区资源优势,依据经济发展水平及“整体、协调、循环、再生”的原则,运用系统工程方法,全面规划,合理组织农业生产,实现农业持续发展和高产高效,达到生态与经济两个系统的良性循环。生态农业建设依据的原理主要包括:
(一)、整体效应原理
这是根据系统观点,即整体功能大于个体功能之和的原理,对整个农业生态系统的结构进行优化设计,利用系统各组分之间的相互作用及反馈机制进行调控,从而提高整个农业生态系统的生产力及稳定性。
(二)、生态位原理(生态位:就是不同物种在同一生活小区内可以占据资源的部位)
各种生物群落在生态系统中都有理想的生态位,在自然生态系统中,随生态演替进行,其生物种群数目增多,生态位丰富并逐渐达到饱和,有利于系统的稳定。另一方面,尽量在农业生态系统中使用不同物种占据不同的生态位,防止生态位重叠造成的竞争互克,使各种生物相安而居。如农田的多层次立体种植、种养结合、水体的立体养殖等,能充分的提高生产效率;
(三)、食物链原理(食物链:表示植食性动物取食植物、肉食性动物取食植食性动物,另一种肉食性动物又取食该种肉食性动物的食物的联系。)
生态农业就是要根据食物链原理组建食物链,将各级营养级上因事物选择所废弃的物质作为营养源,通过混合食物链中相应生物进一步转化利用,使生物能的有效利用率得到提高。
(四)、物质循环与再生原理
任何一个生态系统都有其自身的适应能力与组织能力,可以自我维持和自我调节,而其机制是通过生态系统中物质循环利用和能量流动转化。生态农业讲究尽可能适量或较少的外部投入,通过立体种植及选择归还率较高的作物,以及合理轮作、增施有机肥等建立良性物质循环体系。
(五)、生物种群相生互克原理
自然生态系统中的多种生物种群在其长期进化过程中,形成对自然环境条件特有的适应性,并形成相互依存、相互制约的稳定平衡。我们在生态农业中,专业化利用各种生物种群的相生互克的原理,组建合理高效的复合系统,在有限的空间、时间内容纳更多的生物种,生产更多的产品。
(六)、生物与环境协同进化原理
生物与环境是生态环境的两类组分,也是农业生产的基本要素。生物与环境的协同进化,是指生物在适应环境的同时,也作用于环境,对生态环境有一定的改造能动性,从而使得环境与生物平衡发展。在生态农业中,根据地域生态环境条件,安排生态适应性较好的生物种群,获得较高的生产力水平,并特别注意保护生态环境。
第六节持续农业
一、持续农业的原理:
1.资源环境的持续性:主要是指合理利用资源并使其永继利用,同时防止环境退化.农业资源包括再生资源(光、热、水、土、生物等自然资源)和非再生资源(化肥、农药、机械、水电等)。
2.经济持续性:主要是指农业生产的经济效益及其产品在市场上的竞争能力保持良好和稳定,这直接影响到生产是否能维持和发展下去。
3.社会持续性:指农业生产与国民经济总体发展协调,农产品能满足人民生活水平提高就的需求。
二、持续农业的三大目标
1、保证食物供给的有效性和安全性
2、增加农业收入,扩大农村的就业机会和脱贫致富
3、保护资源环境永继性循环
三、国外持续性农业的实践
美国:“低投入持续农业”和“高效率持续农业”主要是零用种养畜增加有机肥料,以及豆科植物轮作来解决养分供应,减少化肥的投入量,采取综合防治方法控制农田病虫草害,减少农药、除草剂使用,进行品种改良及调整种植制度,以适应低投入技术。
德国:一是强调生态平衡和农业生产系统的良性循环
二是重点防止土壤肥力下降和土壤退化,加强对土地利用、水土流失及病虫防治的管理
三是注意水资源的高效利用,严格控制水源的污染
四是努力降低生产成本,提高农产品在市场上的竞争能力,并且重视生态环境发展与经济发展的关系,加强宏观调控。
日本:“环保型持续农业”
减轻农业对环境造成的负作用,加强对环保型农业技术的研究开发,加强资源的在利用和地力维持与提高等,强调自然资源的循环利用
印度:研制和推广生物肥料,节约化肥施用,推广运用生物农药,减少化学农药,成立农工商企业集团,加强对资源的有效开发和利用及振兴农村经济
中国的“集约持续农业”
1.改善农业生产条件,提高物质投入水平,这是中国农业持续发展的基本保证。
2.努力提高土地生产率,坚持精耕细作的优良传统,高度集约地多维利用土地,提高单位面积产量
3.将传统农业技术与现代农业技术有机结合起来,努力提高各项资源的利用效率和投入效益,并节约和保护资源
4.把提高农业生产的效益和农民收入水平放在重要位置,从宏观上有效加以调控。
5.有效的控制环境退化,尤其要加强农田基本建设,提高抗灾能力,使农业持续发展。把治理与发展协调起来,提高农业综合生产能力,有序高效的利用资源,减轻环境污染和破坏。
第七节生物多样性保护的伦理学基础
对保护生物多样性而言,经济上的考虑是重要的,理论学上的基础也是汇重要的。实际上,更根本的办法是改变我们唯利是图的价值观。特别是那些从经济学的角度看来,一些体型小、种群数量不大、地理分布有限、外表也不吸引人、眼下对人类无用的物种将被赋予低的价值,世界上大部分物种,特别是昆虫、其他无脊椎动物、真菌、细菌和原生动物等均摆脱不了这样的命运。在保护这些物种的投资来讲似乎无任何近起期的经济理由。
因此,伦理学上的考虑将为保护珍稀动物和无明显经济价值的物种提供了充分的理由。
1.每个物种都有存在的权利
2.所有物种是相互依存的
3.人类必须生活在与其他物种相同的生态学限度内
4.人类有责任充当地球的管家,因为人类是自然的一部分,与地球上其它物种一样,受到永恒生态规律的制约。
5.对人类活动和人类多样性的尊重与对生物多样性的兼容,也就是我们要不断成熟达到一个这样的高度,“认同生命存在的所有形式和承认所有物种的固有价值”,从而形成一个扩展的道德义务圈。实现真正的“地球村”
6.自然的精神和美学价值胜过其他的经济价值
7.确定生物起源需要生物多样性
|